
















Abstract
Background. The role of ATPIF1 in ischemic brain injury is rarely reported.
Objectives. This study explored the effect of ATPIF1 on astrocyte activity under oxygen glucose deprivation/reoxygenation (OGD/R).
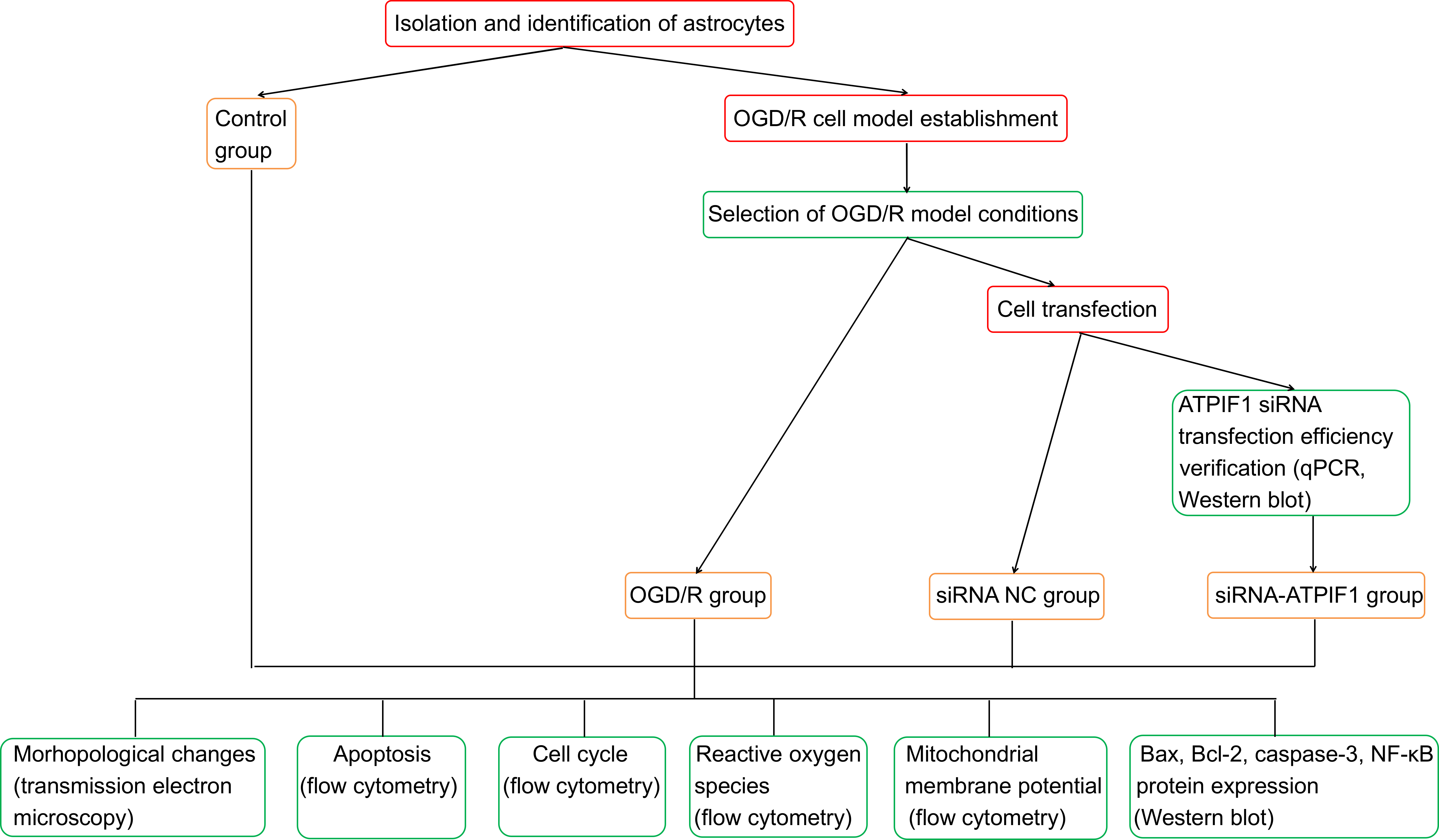
Materials and methods. The study sample was randomly allocated into: 1) control group (blank control); 2) OGD/R group (hypoxia for 6 h/reoxygenation for 1 h); 3) siRNA negative control (NC) group (OGD/R model+siRNA NC); and 4) siRNA-ATPIF1 group (OGD/R model+siRNA-ATPIF1). The OGD/R cell model was established from Sprague Dawley (SD) rats to simulate ischemia/reperfusion injury. Cells in the siRNA-ATPIF1 group were treated with siATPIF1. Ultrastructural changes in the mitochondria were observed using transmission electron microscopy (TEM). Apoptosis, cell cycle, reactive oxygen species (ROS), and mitochondrial membrane potential (MMP) were detected with flow cytometry. The protein expression levels of nuclear factor kappa B (NF-κB), B-cell lymphoma 2 (Bcl-2), Bcl-2-associated X (Bax), and caspase-3 were detected with western blot.
Results. In the model group, the cell structure and the ridge structure were destroyed, and mitochondria edema, outer membrane damage and vacuole-like lesions were observed. Compared with the control group, the OGD/R group had considerably increased apoptosis, G0/G1 phase, ROS content, MMP, and Bax, caspase-3 and NF-κB protein expression, as well as markedly decreased S phase and Bcl-2 protein expression. Compared with the OGD/R group, the siRNA-ATPIF1 group had considerably decreased apoptosis, G0/G1 phase, ROS content, MMP, and Bax, caspase-3 and NF-κB protein expression, as well as remarkably increased S phase and Bcl-2 protein expression.
Conclusions. The inhibition of ATPIF1 may alleviate OGD/R-induced astrocyte injury by regulating the NF-κB signaling pathway, inhibiting apoptosis, and reducing the ROS content and MMP in the rat brain ischemic model.
Key words: apoptosis, NF-κB signaling pathway, OGD/R, astrocytes, ATPIF1
Background
Ischemic cerebrovascular disease has been the leading cause of disability and mortality globally.1, 2 A variety of biochemical mechanisms are involved in the pathophysiological process of the disease, including brain energy metabolism disorder, reactive oxygen species (ROS) production, inflammation, acidosis, and apoptosis.3 As the most widely distributed and numerous cells in the brain, astrocytes are very important for maintaining neuronal structure and function.4, 5 When ischemic injury occurs, astrocytes can fill the damaged area, maintain the stability of the environment in the central nervous system, provide nutrients for neurons, and protect neurons; thus, they are crucial for the repair of an ischemic area and to avoid the secondary injury of neurons.6
All cell activities are inseparable from adenosine triphosphate (ATP) consumption; hence, ATP synthesis and decomposition in astrocytes are of great importance after the ischemic injury. Mitochondrial ATPase inhibitor, ATPIF1, is a nuclear coding protein that can interact with ATP synthase.7 The ATPIF1 is the most characteristic regulator of F1Fo-ATP synthase. It combines with enzymes inhibiting its hydrolytic activity to protect cells from ATP consumption.8 The ATPIF1 has been associated with a variety of diseases, such as diabetic nephropathy, breast cancer and anemia.9, 10, 11, 12 The ATPIF1 can aggravate the development of diabetic nephropathy by inducing mitochondrial damage.9 The inhibition of ATPIF1 can improve the severe decrease in electron transport chain activity in mitochondrial pathology.10 However, the role of ATPIF1 in ischemic brain injury is rarely reported.
In biological systems, the mitochondria are the main place for cells to generate energy. They are also the key intracellular targets for initiating the intrinsic pathway of apoptosis.13 The ATPIF1 abnormality causes a series of chain reactions in the mitochondria. The mitochondria are the main part of ROS production in tissues. Increased ROS levels can induce apoptosis through endogenous and exogenous pathways.14 Nuclear factor kappa B (NF-κB) is a traditional transcription factor. It regulates the apoptosis program through its downstream target genes under different stimuli, such as ROS accumulation and inflammatory factors.15 However, the role of ATPIF1 in astrocyte apoptosis mediated by oxygen glucose deprivation/reoxygenation (OGD/R) remains unknown.
Objectives
The aim of this study was to explore the effect of ATPIF1 on astrocyte activity in Sprague Dawley (SD) rats under OGD/R. We hypothesized that siATPIF1 has a protective effect on the astrocyte activity in OGD/R-induced astrocyte injury. The findings of this study may help develop a feasible approach to treating ischemic cerebrovascular disease.
Materials and methods
Ethics declarations
The Animal Care and Use Committee of the Affiliated Hospital of Zunyi Medical University, China, has given the approval to the experiments and protocol used in this study (approval No. 2020041501).
Experimental animals
Specific pathogen-free SD rats (female, 3 months old; male, 9 months old; license No. SCXK (Xiang) 2019-0014) were obtained from Changsha Tianqin Biotechnology Co., Ltd. (Hunan, China) and raised under 30–70% humidity, 20–26°C and 12 h/12 h light/dark cycle. The rats were allowed food and water ad libitum.
Primary cell isolation and identification
The rats were caged together with a male–female ratio of 1:2, and the neonatal rats were used for further experiments. No gender requirement was set for the neonatal rats. A total of 4 litters with about 12 neonatal rats in each litter were used. The neonatal rats were given sodium pentobarbital (45 mg/kg; MilliporeSigma, Burlington, USA). The brains of the rats were removed following cervical dislocation after immersion and disinfection with 75% ethanol for 1 min. The pia mater covered on the surface of the brain tissue was stripped, and the rat cerebral cortex was separated and washed repeatedly with D-Hank’s solution (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China). The washed cortex was repeatedly ground and sieved (220 mesh screen). The screen was repeatedly washed and the cell suspension was transferred to a 15-milliliter clean tube. The pooled samples were centrifuged at 4°C (1500 rpm) for 5 min. The supernatant was discarded, the precipitate was resuspended in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Thermo Fisher Scientific, Waltham, USA) containing 10% fetal bovine serum (FBS; Beijing Solarbio Science & Technology Co., Ltd.), and the cells were cultured in 5% CO2 at 37°C. The medium was replaced with a fresh medium every 3 days. When the cells reached 90% confluence, they were placed in a 37°C shaker incubator for 4–6 h. Then, the microglia supernatant was removed.
The cell climbing sheets were washed with 0.1 M phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde (Beijing Solarbio Science & Technology Co., Ltd.) at room temperature (RT) for 15 min, and blocked for 40 min with 3% goat serum (Beijing Solarbio Science & Technology Co., Ltd.) and 0.1% Triton X-100 (MilliporeSigma). The cells were incubated with rabbit anti-glial fibrillary acidic protein (anti-GFAP) primary antibody (1:200, AB53554; Abcam, Cambridge, UK) at 4°C overnight and IgG/Cy3 fluorescent secondary antibody (1:200, A0502; Beyotime Biotechnology Inc., Shanghai, China) at RT for 60 min. Then, 4’,6-diamidino-2-phenylindole (DAPI; Jiangsu KeyGen Biotech Corp., Ltd., Jiangsu, China) was applied and incubated for 3 min. The cells were mounted with 95% glycerol and observed using a fluorescence microscope (model CX41; Olympus Corp., Tokyo, Japan).
OGD/R cell model
Third-generation astrocytes with good growth condition were selected and 0.125% trypsin (Beijing Solarbio Science & Technology Co., Ltd.) was added for digestion. The cells were centrifuged at 1500 rpm for 5 min and the precipitation was resuspended. The cells were seeded in a 6-well culture plate (1×105/well) and cultured in sugar-free DMEM with 94% N2, 5% CO2 and 1% O2 at 37°C for 24 h. The cells were collected and detected at 1, 3, 6, and 24 h.
Cell transfection
The cells were cultured in a 6-well plate. Cell transfection was performed when they reached 70% confluence. Lipofectamine 3000 kit (Invitrogen, Thermo Fisher Scientific, Waltham, USA) was used. A 12.5 μL small interfering RNA (siRNA) was mixed with 125 μL of Opti-MEM (Gibco) and 5 μL of Lipofectamine 3000 was mixed with 125 μL of Opti-MEM. Both mixtures were separately incubated at RT for 5 min, then mixed, incubated for 15 min at RT, and added to the cells cultured in the 6-well plate for 4 h. The cells were cultured in DMEM/F-12 complete medium (Gibco) containing 20% FBS (BBI Life Sciences Corp., Shanghai, China).
The transcription sequences were obtained from the National Center for Biotechnology Information (NCBI; https://www.ncbi.nlm.nih.gov/). The target gene sequences were handed over to General Biol Co., Ltd. (Anhui, China), which directly provided the siRNA. The siRNA sequences are shown in Table 1.
Experimental grouping
The experimental grouping was randomly divided into:
1) control group (blank control);
2) OGD/R group (hypoxia for 6 h/reoxygenation for 1 h);
3) siRNA negative control (NC) group (OGD/R model +siRNA NC);
4) siRNA-ATPIF1 group (OGD/R model+siRNA-ATPIF1).
The investigators performing the subsequent experiments were blinded to the group allocation.
Quantitative real-time polymerase
chain reaction
The cells were collected and Trizol reagent (CWBIO, Jiangsu, China) was used for the total RNA extraction. Reverse transcription to cDNA was performed with a HiFiScript cDNA Synthesis Kit (Vazyme Biotech, Jiangsu, China) according to the kit instructions. Quantitative PCR was conducted with a quantitative real-time polymerase chain reaction (qPCR) system (CFX Connect; Bio-Rad, Hercules, USA). The reaction system and reaction steps are presented in Table 2. The primers are shown in Table 3. The β-actin (Sino Biological Inc., Beijing, China) served as the internal control. The relative ATPIF1 mRNA expression was determined based on the 2−ΔΔCt method.
Western blot
The cells were lysed in the radioimmunoprecipitation (RIPA) buffer (Applygen Technologies Inc., Beijing, China) at 4°C for 30 min and then centrifuged at 12,000 rpm for 10 min. For total protein determination, the supernatant was carefully extracted, and determination was conducted according to the Bradford method (Beijing Solarbio Science & Technology Co., Ltd.). This step was followed by protein denaturation and sample loading. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE; Xilong Scientific Co., Ltd., Guangdong, China) was performed for 2 h before transferring to polyvinylidene fluoride membrane (PVDF; MilliporeSigma) at 300 mA. The primary antibodies used included mouse monoclonal anti-glyceraldehyde 3-phosphate dehydrogenase (anti- GAPDH; 1:2000, cat. No. TA-08; ZSGB-BIO, Beijing, China), mouse anti-ATPIF1 (1:1000, ab110277; Abcam), mouse anti-B-cell lymphoma 2 (anti-Bcl-2; 1:1000, cat. No. 60178-1-Ig; Proteintech, Rosemont, USA), rabbit anti-Bcl-2-associated X (anti-Bax; 1:1000, cat. No. 50599-2-Ig; Proteintech), rabbit anti-nuclear factor-kappa B (anti- NF-κB; 1:1000, 10745-1-AP; Proteintech), and rabbit anti-caspase-3 (1:1000, cat. No. YT0656; ImmunoWay Biotechnology Co., Plano, USA). The primary antibodies were added and incubated at 4°C overnight. Then, the secondary antibodies were added and incubated at RT for 2 h. The secondary antibodies used included goat anti-rabbit IgG antibody (H+L, horseradish peroxidase (HRP)-conjugated, 1:2000, cat. No. ZB-2301) and goat anti-mouse IgG antibody (H+L, HRP-conjugated, 1:2000, cat. No. ZB-2305; ZSGB-BIO). The chemiluminescent substrate was added, and the membrane was examined using a gel imaging system (ChemiDoc XRS+; Bio-Rad). The blot was analyzed using Quantity One software v. 4.62 (Bio-Rad) with GAPDH as the internal reference, using the equation: relative protein expression = (target protein integral optical density (IOD) / internal reference protein IOD).
Detection with flow cytometry
Apoptosis
The cells (1×106–3×106) were collected. An Annexin V–Propidium Iodide (PI) Analysis Kit (MultiSciences (Lianke) Biotech Co., Ltd., Hangzhou, China) was used for apoptosis detection based on the manufacturer’s instructions. The cells were washed 3 times with pre-cooled PBS, resuspended with 300 μL of ×1 binding buffer, added with 3 μL of Annexin V-fluorescein isothiocyanate (FITC) and 5 μL of PI-phycoerythrin solution, and mixed gently. Then, the cells were incubated in the dark at RT (20–25°C) for 10–20 min and placed in an ice bath. The detection was performed on a NovoCyte™ flow cytometer (NovoCyte 2060R; ACEA Bio (Hangzhou) Co., Ltd., Hangzhou, China).
Cell cycle
The cells were collected, washed with PBS, 1 mL of DNA staining solution and 10 μL of PI solution were added, and the cells were mixed. Then, they were incubated in the dark at RT for 30 min. The cell cycle was determined with flow cytometry.
ROS
The cells were harvested and a final concentration of 10 μM 2’,7’-dichlorofluorescein diacetate (DCFH-DA) solution (Reactive Oxygen Species Assay Kit; Jiangsu KeyGEN BioTECH Co., Ltd.) was obtained with1:1000 dilution of DCFH-DA in serum-free medium. The cell culture medium was replaced with the diluted DCFH-DA. The cells were incubated at 37°C for 20 min and then washed 3 times with serum-free medium and 3 times with PBS. The ROS levels were determined with flow cytometry.
Membrane potential
The cells were collected, washed with PBS and centrifuged at 2000 rpm for 5 min. Next, the cells were resuspended with 500 µL of JC-1 solution (BestBio, Shanghai, China) and cultured at 37°C with 5% CO2 for 15–20 min. Then, they were centrifuged at 2000 rpm for 5 min, washed twice with ×1 incubation buffer, resuspended with 500 µL of ×1 incubation buffer, and analyzed with flow cytometry.
Statistical analyses
Data analysis was performed with GraphPad Prism 7 software (GraphPad Software, San Diego, USA). The comparisons between groups were performed using one-way analysis of variance (ANOVA) with Bonferroni post hoc test. The value of p < 0.05 was considered statistically significant.
Outcome measures
The outcome measures were the effects of siATPIF1 on the mitochondrial structure, apoptosis, cell cycle, ROS production, mitochondrial membrane potential (MMP), apoptosis-related genes, and the NF-κB pathway in the OGD/R model.
Results
Isolation and identification of astrocytes
The isolated and purified cells were star-shaped, and many long and branched processes were separated from the cell body. The GFAP is a marker for astrocytes. Immunofluorescence results showed that the nuclei of astrocytes were stained blue and the cells were stained red (GFAP). The purity of astrocytes was more than 95% (Figure 1).
Selection of OGD/R model conditions
As shown in Figure 2A, the apoptosis rate was the highest at 1 h of reperfusion compared with the control group. Many cells were irregular and had mitochondrial edema, outer membrane damage, vacuole-like lesions, or damaged ridge structure at 1 h of reperfusion (Figure 2B). Moreover, the mitochondria were irregular, some mitochondria were swollen, and the cristae disappeared at 3 h after reperfusion. Finally, the mitochondrial ridge structure was destroyed and became a vacuole-like lesion at 24 h after reperfusion. Figure 2C shows that ATPIF1 protein expression increased remarkably at 1 h of reperfusion. Therefore, the model was established at 1 h of reperfusion in subsequent experiments.
ATPIF1 siRNA transfection efficiency verification
As shown in Figure 3, the ATPIF1 mRNA expression levels in the siRNA groups (particularly the siATPIF1-2 group) were considerably lower than those in the control and siRNA NC groups. The ATPIF1 protein expression levels in the siRNA-2 and siRNA-3 groups were remarkably lower than in the control and siRNA NC groups. Therefore, siRNA-2 was selected for subsequent experiments.
Effect of siATPIF1 on mitochondrial structure in the OGD/R model
The mitochondrial morphological changes were examined using transmission electron microscopy (TEM). As shown in Figure 4, the cell morphology in the control group was normal and the ridge structure was complete; the cell and ridge structures in the OGD/R group were destroyed, and mitochondria edema, outer membrane damage or vacuole-like lesions were noted; the cells in the siRNA NC group were irregular, some mitochondria were swollen and cristae disappeared; the cell and mitochondrial membrane structures in the siRNA-ATPIF1 group were complete, and most of the ridge structures were normal.
Effects of siATPIF1 on apoptosis
and cell cycle in the OGD/R model
The apoptosis rate in the OGD/R group was considerably higher than in the control group, whereas the apoptosis rate in the siRNA-ATPIF1 group was substantially lower than in the OGD/R group (Figure 5A). The OGD/R group had a remarkably increased G0/G1 and a remarkably decreased S phase compared with the control group. Moreover, the siRNA-ATPIF1 group had a considerably decreased G0/G1 phase and substantially increased S phase compared with the OGD/R group (Figure 5B).
Effects of siATPIF1 on ROS and MMP in the OGD/R model
The ROS levels in the OGD/R group were significantly increased compared with the control group; the ROS levels in the siRNA-ATPIF1 group were significantly decreased compared with the OGD/R group (Figure 6A). The MMP in the OGD/R group was considerably increased compared with the control group, and the MMP in the siRNA-ATPIF1 group was remarkably decreased compared with the OGD/R group (Figure 6B).
Effects of siATPIF1 on apoptosis-related genes and NF-κB pathway in the OGD/R model
The western blot results are shown in Figure 7. The OGD/R group had substantially increased Bax, caspase-3 and NF-κB protein expression, and remarkably decreased Bcl-2 protein expression compared with the control group. The siRNA-ATPIF1 group had remarkably decreased Bax, caspase-3 and NF-κB protein expression levels, and increased Bcl-2 protein expression compared with the OGD/R group.
Discussion
Evidence of the relationship between ischemic brain injury and increased astrocyte injury has been found in previous research.16 Astrocytes are the most abundant cell type in the central nervous system.17 They communicate with neurons, oligodendrocytes and endothelial cells, and participate in maintaining normal brain function.18 Therefore, astrocytes have become the neuroprotective target in ischemic brain injury.19 The ATPIF1 is a mitochondrion-localized protein encoded by nuclear DNA. The ATPIF1 can inhibit ATP hydrolysis through ATP synthase when bound to F1 complexes α and β.20 The ATPIF1 regulates mitochondrial morphology and cell survival.21 Our study demonstrated that siRNA-ATPIF1 may protect astrocytes from OGD/R-induced apoptosis and necrosis by maintaining mitochondrial anti-apoptosis (Bcl-2 upregulation) and bioenergetic function.
Apoptosis can be induced by extrinsic (death receptor) and intrinsic (mitochondrial) pathways.22 Mitochondrial morphology changes in the mitochondria-mediated pathway. The increase in mitochondrial membrane permeability causes the activation of downstream effector caspases, such as caspase-3, and eventually induces apoptosis.23 Xu et al. demonstrated that ATPIF1 plays a key role in regulating cleaved caspase-3 expression in the microglia.24 The Bcl-2 family proteins, such as Bax and Bcl-2, mediate the intrinsic pathway by altering the outer membrane permeability of the mitochondria.25 The Bax protein is a common channel of the mitochondria-mediated apoptosis pathway.26 We found that siRNA-ATPIF1 could inhibit Bax and increase Bcl-2 protein expression in astrocytes. The OGD/R-induced astrocytes had increased MMP and abnormal mitochondrial morphology, and siRNA-ATPIF1 inhibited these processes. Cleaved caspase-3 expression decreased in the OGD/R cells treated with siRNA-ATPIF1. The results showed that siRNA-ATPIF1 could effectively inhibit mitochondria-mediated apoptosis in astrocytes after the OGD/R treatment.
The NF-κB plays an active role in apoptosis.27 In a nonstimulated state, NF-κB and IκB form a complex in the cytoplasm, but they are not activated. The IκB is rapidly phosphorylated after cell stimulation and further degraded by ubiquitinase or protease.28 After IκB degradation is complete, NF-κB is rapidly activated and then translocated into the nucleus to react with the downstream targets.29 The NF-κB can respond to ROS accumulation signals and initiate the pro-apoptotic pathway activation.30 The present study found that astrocytes produce a large amount of ROS after OGD/R. Therefore, we investigated whether NF-κB is involved in ATPIF1-mediated mitochondrial apoptosis. Western blot results showed that ATPIF1 knockdown could reduce NF-κB expression, indicating that the NF-κB signaling pathway may be involved in ATPIF1-induced mitochondrial apoptosis in astrocytes.
Limitations
Although this study has clarified the effect of ATPIF1 on astrocyte apoptosis, some studies have pointed out that pH changes also have a remarkable effect on ATPIF activity.31 However, this factor was not taken into account in our experiments. The effect of hypoxia on pH value in vivo and then on ATPIF1, particularly in animals and even in humans, will also be a direction of our subsequent studies.
Recently, the application of ATPIF1 gene-silenced T cells in the preparation of antitumor drugs has also been described, which provides a new direction for the research and development of adoptive immunotherapy for tumors. We believe that cell therapy by ATPIF1 gene knockout can have a broad development prospect in the treatment of cerebral ischemia injury.
Conclusions
This study is the first to report that during ischemic brain injury, ATPIF1 regulation of mitochondria-mediated apoptosis is associated with the promotion of NF-κB activation. The ATPIF1 knockdown can improve the mitochondrial morphology, inhibit MMP, massive ROS accumulation and NF-κB activation, reduce Bax expression, increase Bcl-2 protein expression, and maintain astrocyte viability in OGD/R-induced injury.
Generalizability/translation
The findings of this study may be generalized to other species or experimental conditions, including human biology.
Data availability statement
Data will be made available by the corresponding author upon reasonable request.
Supplementary files
The supplementary files are available at https://doi.org/
10.5281/zenodo.7454344. The package contains the following files:
Supplementary Table 1. Data analysis for Figure 2, Figure 3, Figure 5, Figure 6, Figure 7.






