
















Abstract
Background. Hypertensive disorders complicating pregnancy (HDCP) are one of the most serious medical disorders during pregnancy.
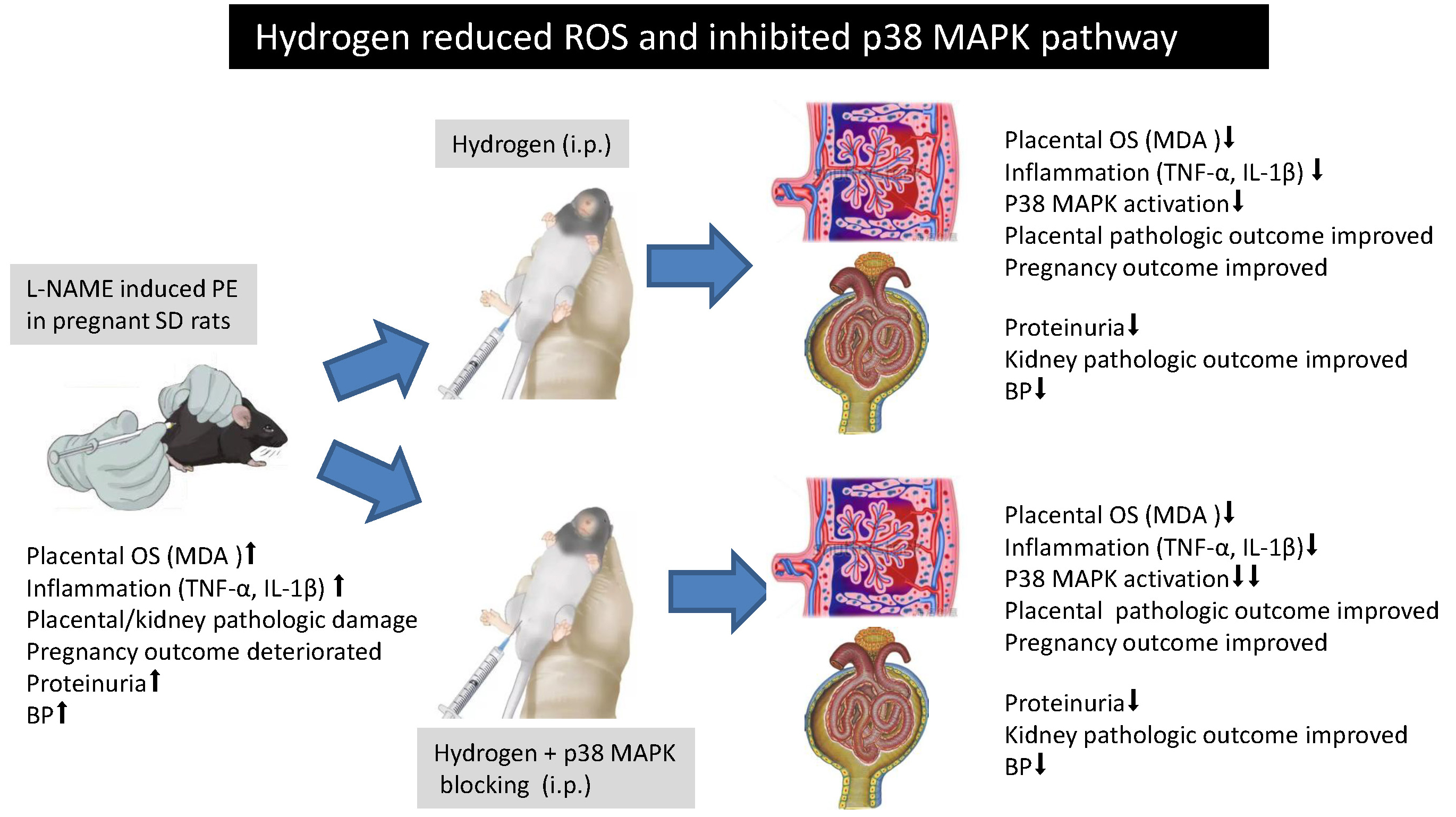
Objectives. To investigate the effects of hydrogen on the mitogen-activated protein kinase (MAPK) signaling pathway in preeclampsia (PE).
Materials and methods. The N(omega)-nitro-L-arginine methyl ester (L-NAME)-induced PE model with Sprague Dawley (SD) rats was employed. An inhibitor of MAPK signaling pathways (SB203580) was used as a p38 MAPK inhibitor. The SD rats were randomized into 5 groups: non-pregnant (NP); normal pregnancy (P); pregnancy + L-NAME (L); pregnancy + L-NAME + hydrogen-rich saline (LH); and pregnancy + L-NAME + hydrogen-rich saline + SB203580 (LHS). The pregnancies were terminated on day 22 of gestation, and the placentas and kidneys were microscopically inspected. Tumor necrosis factor alpha (TNF-α), interleukin-1β (IL-1β) and malondialdehyde (MDA) levels were assessed. The mean systolic blood pressure (SBP) and level of proteinuria were recorded. The p38 MAPK mRNA expression and p-p38 MAPK protein levels were measured using real-time polymerase chain reaction (RT-PCR) and western blot, respectively.
Results. It was found that hydrogen-rich saline (LH group) decreased placental MDA, proteinuria, TNF-α, and IL-1β levels in the placental tissues compared with the L group (all p < 0.05). Additionally, hydrogen-rich saline (LH group) treatment significantly decreased the p38 MAPK mRNA expression and p-p38 MAPK protein levels compared with the L group (p < 0.05). The p38 MAPK inhibitor SB203580 (LHS group) further decreased the p38 MAPK mRNA expression and p-p38 MAPK protein levels compared with the LH group (p < 0.05).
Conclusions. Hydrogen can decrease the reactive oxygen species (ROS) content and inhibit the MAPK pathway. The protective effect of hydrogen may be associated with the inhibition of the p38 MAPK signaling pathway.
Key words: preeclampsia, antioxidant, reactive oxygen species, hydrogen, p38 MAPK pathway
Background
Preeclampsia (PE) is a serious disease for pregnant women. It can lead to placental disorders, fetal growth restriction and other unfavorable outcomes.1 Its etiology remains complex.
The presence of oxidative stress, such as placental oxidative stress2 or periodontal oxidative stress,3 may increase the risk of PE in pregnant women. As a novel antioxidant, hydrogen selectively reduced toxic reactive oxygen levels,4 which was observed in a PE rat model in our previous study,5 indicating that hydrogen is a potential therapeutic antioxidant that could be clinically applied in treating PE. Its mechanism of action remains to be investigated.
Previous studies found that oxidative stress activated mitogen-activated protein kinase (MAPK) signaling and increased the expression of apoptosis receptors.6 The p38 MAPK activity was observed to be significantly higher in women with PE placentas.7 These findings imply that reactive oxygen species (ROS) could activate the p38 MAPK signaling pathway in the placenta, followed by the overexpression of sFlt-1 and sEng in maternal serum8 and a systemic inflammatory reaction, giving rise to endothelial dysfunction and the clinical symptoms of PE. Therefore, hydrogen, as an antioxidant agent, may have an effect on the MAPK signaling pathway by blocking ROS, thereby reducing the inflammatory reaction and apoptosis, and ultimately improving the prognosis of PE patients.
To explore the mechanism of molecular hydrogen in the pathogenesis of hypertensive disorders complicating pregnancy (HDCP), we investigated the influence of hydrogen on ROS content, apoptosis and the MAPK signaling pathway at the mRNA and protein levels by employing a previously established N(omega)-nitro-L-arginine methyl ester (L-NAME)-induced gestational hypertension rat model.
To further confirm the role of the MAPK signaling pathways in the pathogenesis of HDCP, an inhibitor of MAPK signaling pathways (SB203580) was employed in the L-NAME-induced gestational hypertension rat model.
The investigation of the mechanism of molecular hydrogen could potentially promote novel treatment strategies in the PE and HDCP patient populations.
Objectives
The objective of this study was to explore the mechanism of molecular hydrogen in the pathogenesis of PE by investigating the influence of hydrogen on ROS content, apoptosis and the MAPK signaling pathway at the mRNA and protein levels. We employed a previously established gestational hypertension rat model5 and an inhibitor of MAPK signaling pathways (SB203580).9
Materials and methods
The experiment was approved by animal ethical committee of Tongji University, Shanghai, China (approval No. TJLAC-020-010).
Animals and drugs
Fifty female and 50 male Sprague Dawley (SD) rats weighing 220–250 g and aged 9–10 weeks were employed. They were kept under light-, air- and temperature-controlled conditions for 7 days before the experiment. The rats were mated separately overnight. The first day of finding spermatozoa using vaginal smear in the female rat was defined as day 0 of gestation for the female rat. Each pregnant rat was raised in a separate metabolic cage. On the 21st gestational day, the metabolic cages were used to collect 24-hour urine samples.
Hydrogen was dissolved in physiological saline for 6 h under 0.4 MPa, followed by sterilization. The prepared hydrogen saline was stored in an aluminum bag at 4°C. Gas chromatography was used to measure the concentration of hydrogen.4 Hydrogen saline above 0.6 mmol/L, L-NAME and SB203580 (both from Sigma-Aldrich, St. Louis, USA) were used in the experiment.
Intervention
The rats were randomized into 5 groups on the 15th day of pregnancy: non-pregnant (NP); normal pregnancy (P); pregnancy + L-NAME (L); pregnancy + L-NAME + hydrogen-rich saline (LH); and pregnancy + L-NAME + hydrogen-rich saline + SB203580 (LHS). Each group had 10 rats. The NP rats and P rats as control groups received an intraperitoneal injection of normal saline (5 mL/kg). The PE group (L) and hydrogen intervention group (LH) received L-NAME at a dose of 60 mg/kg dissolved in normal saline or hydrogen-rich saline, respectively. In the LHS group, the pregnant rats received 10 mg/kg/d of SB203580 1 h before receiving L-NAME at a dose of 60 mg/kg dissolved in hydrogen-rich saline.
All rats were anesthetized with ether on the 22nd day of pregnancy. The systolic blood pressure (SBP) of the right carotid artery was measured with a polyethylene catheter connected to a Medlab pressure transducer (Nanjing Biotech Co., Jiangsu, China). All rats were sacrificed on the same day under ketamine anesthesia. The pregnant rats underwent cesarean section. The resorption and weight of each pup were determined.
Partial tissues of the placenta, kidney, liver, and aorta were fixed with paraformaldehyde (40 g/L), and the remaining tissues were stored in liquid nitrogen for a subsequent analysis.
Biochemical factors and cytokines
Following the manufacturer’s instructions for the malondialdehyde (MDA) kit (Nanjing Biotech Co.), placental MDA was measured in 100 mg of wet tissue. Twenty-four-hour urine samples were collected and quantified with a total protein kit (Sigma-Aldrich). Placental cytokines, including tumor necrosis factor alpha (TNF-α) and interleukin-1β (IL-1β), were measured using an enzyme-linked immunosorbent assay (ELISA) kit (Biosource International, Inc., Camarillo, USA). Based on a standard curve, the TNF-α and IL-1β concentrations were calculated using an absorbance reader (Denley Dragon; Thermo Fisher Scientific, Helsinki, Finland).
Real-time polymerase chain reaction
Total RNA was extracted from tissues preserved in liquid nitrogen with TRIpure Total RNA Extraction Reagent (ELK Biotechnology Co., Wuhan, China). The EntiLink™ Synthesis Kit (EQ003; ELK Biotechnology) was employed to synthesize the first strand of cDNA, followed by real-time fluorescence quantitative polymerase chain reaction (RT-PCR) with the EnTurbo™ SYBR Green PCR SuperMax Kit (EQ001; ELK Biotechnology), which was performed on a StepOne™ Real-Time PCR Instrument (Life Technologies, Carlsbad, USA). The comparative CT method (ΔΔCT) was used for relative gene expression analysis. All PCR reactions were performed in duplicate. Primer Premier v. 5.0 software (PREMIER Biosoft International Co., San Francisco, USA) was used for the design of the primer sequences.
Western blot
Western blot was performed as previously described.10 The total protein of the placental tissue was extracted using whole cell lysis buffer. The protein samples were resolved on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) gels (Beyotime Biotechnology, Shanghai, China). The proteins were transferred to polyvinylidene fluoride (PVDF) membranes. Rabbit anti-rat primary antibody was probed on the membranes after blocking. Using the ECL detection system (GeneCopoeia Inc., Rockville, USA), we determined the immune complexes by incubation with a secondary antibody, which was conjugated with horseradish peroxidase (HRP).
Statistical analyses
The SPSS v. 13.0 (SPSS Inc., Chicago, USA) and GraphPad v. 6.0 (GraphPad Software, San Diego, USA) software was used for statistical analysis. Data distribution was analyzed using the Shapiro–Wilk method. The Student’s t-test was used for comparison between the 2 groups for normally distributed data, and the Mann–Whitney U test was used for non-normally distributed data between the 2 groups. The Levene’s test was used to determine the homogeneity of variance for the normally distributed data. Statistical significance was defined as p ≤ 0.05. All statistical data can be found in the Supplementary files (https://doi.org/10.5281/zenodo.7074902).
Results
Basic parameters
The number of resorptions, fetal weight, protein excretion, and SBP are shown in Table 1. Administration of L-NAME (L group) caused a significant increase in the number of resorptions, SBP and 24-hour urine protein excretion, as well as a decrease in fetal weight, compared with the normal pregnant rats (P group). The hydrogen intervention resulted in an improvement in pregnancy outcomes in terms of fetal weight, resorptions, urine protein excretion, and SBP. However, there were no differences in all indices between the LH and LHS group.
Lipid peroxidation product levels
The L-NAME-treated group (L group) showed a marked increase in the level of placental MDA on the terminal day (p < 0.001; Figure 1), while the hydrogen-treated rats showed a significant decrease of MDA in both the LH and LHS groups when compared with L group, respectively (p < 0.001). Malondialdehyde levels were lower in the P group than in the other groups (p < 0.001).
Inflammatory factor levels
The L-NAME-treated group (L group) showed elevated placental TNF-α and IL-1β levels, which was in contrast with the P group (both p < 0.001, Table 2). However, the hydrogen intervention considerably depressed the L-NAME-mediated increase in placental TNF-α and IL-1β levels. The LHS group showed an even further decrease of inflammatory factor levels compared with the LH group (p < 0.001 and p = 0.002, respectively).
Histopathology outcome
When compared with NP and P groups (Figure 2A,B), reduced Bowmen’s capsule space and hyperplasia in the glomerular epithelium, in addition to degenerative changes in the proximal convoluted tubules, were significant in the L group (Figure 2C). This result was consistent with the elevated urine protein levels caused by the administration of L-NAME. Interestingly, the pathological changes were markedly alleviated in the LH and LHS groups (Figure 2D,E). Similarly, abnormalities were absent in both the NP and P control groups (Figure 2A,B). Compared with the P group (Figure 3A), L-NAME caused a remarkable degeneration in the junctional zone of placenta (Figure 3B), but such degeneration was not found in the P, LH and LHS groups (Figure 3A,C,D). Similarly, the result of apoptosis in the L group was consistent with findings from the hematoxylin and eosin (H&E)-stained specimens (p < 0.001, Figure 4, Figure 5). When compared with NP and P groups (Figure 6A,B), the continuous structure of the hepatic lobules was interrupted, accompanied by hepatocyte edema in the L group (Figure 6C). Moderate edema was found in the LH and LHS groups (Figure 6D,E). No significant differences were observed in the aorta between the groups (Figure 7).
Real-time PCR outcome
The L-NAME intervention in the L group caused higher p38 mRNA expression when compared with the P group (p < 0.001, Figure 8). The p38 mRNA in the placenta was depressed in LH and LHS groups (p < 0.001). We also noticed that the p38 MAPK inhibitor SB203580 (LHS group) decreased p38 MAPK mRNA expression more than the hydrogen-only intervention (LH group, p < 0.001). The p38 MAPK mRNA expression did not differ significantly between P and LHS groups.
Western blot outcome
Group L showed an elevation of placental p-p38 MAPK protein level compared to the P group (p < 0.001, Figure 9). Lower p-p38 MAPK protein levels in placental tissues were found in the LH and LHS groups compared to the L group (p < 0.001). The p38 MAPK inhibitor SB203580 (LHS group) further decreased the p-p38 MAPK protein level compared with the hydrogen-only intervention (LH group, p < 0.001).
Discussion
Abnormal invasion of the spiral arteries11 in PE patients was shown to result in blood resistance changes,12 placental oxidative stress and elevated ROS generation.12, 13 As a “second messenger” in intracellular signaling cascades, ROS influenced cell apoptosis or proliferation14 and played an important role in regulating cellular signaling.15 Among the potential cellular signaling pathways that could be impacted by ROS, the p38 MAPK pathway is a crucial candidate; it was shown to conduct signal transduction from the cell surface to the nucleus,16, 17, 18 which would affect cellular survival, migration, proliferation, and differentiation.19, 20, 21 Accumulated evidence indicates that the p38 MAPK pathway is one of the key signal pathways related to PE pathogenesis.8, 19, 20 It has been demonstrated that nitration of phospho-p38 MAPK was higher in PE placentas.22 Reduced phospho-p38 MAPK concentration and p38 MAPK catalytic activity were also found in placentas from PE cases.22 Trophoblastic p38 signaling may be activated by hypoxic and ischemic stress, thus impacting villous trophoblastic functions in the placentas of PE patients.23 Due to hypoxic stress generated by ROS, the p38-Wip1 feedback regulatory loop is dysregulated in PE patients.20
In this study, we confirmed the role of p38 MAPK in the pathogenesis of PE by employing a rat model. We found that p38 mRNA expression and p-p38 MAPK protein levels in the placenta were significantly elevated in the L-NAME-induced PE rats, which could be associated with increased ROS levels, as indicated by higher MDA. Our findings are consistent with clinical evidence. First, p38 MAPK activity was observed to be significantly higher in women with PE placentas.7 Second, p38 MAPK signaling could be activated through oxidative stress and increased apoptosis receptor expression.6
Interestingly, the expression of p38 was upregulated in the upper uterine segment of pregnant non-laboring or spontaneously laboring uteri, but uniformly distributed in non-pregnant uteri.24 We proposed a hypothesis that placental oxidative stress is elevated in normal pregnancy compared with non-pregnancy status. The p38 expression change in pregnancy could be self-adaption to placental oxidative stress.
As dysregulated TNF-α synthesis may be a consequence of the p38 MAPK activation,25 we also observed abnormal elevation of inflammatory cytokines, including TNF-α and IL-1β in the placenta of the rat model, which may be due to the activation of p38 MAPK cascades. In addition, the p38 MAPK pathway has been regarded as one of the key pathways for a significant decline of microvessel density8 and excessive apoptosis20 in placentas. Similarly, we found that remarkable degeneration and excessive apoptosis in the placentas of the PE rat model and adverse pregnancy outcomes were associated with p38 MAPK signal activation. Furthermore, the activation of the p38 MAPK pathway in PE followed by placental nicotinamide adenine dinucleotide phosphate (NADPH) oxidase19 induced elevated sFlt-l and sEng expression in the maternal circulation.8 This led to systemic vascular endothelial cell dysfunction8 and the pathogenesis of PE.26 In the present study, the renal dysfunction (indicated by 24-hour urine protein excretion) and renal pathological degeneration may result from systemic vascular endothelial cell dysfunction related to p38 MAPK signal activation.
We also investigated the mechanism of action of hydrogen, a novel selective antioxidant. Although various antioxidants used to balance ROS, including vitamin E and vitamin C, have been investigated, the clinical outcomes are still controversial. Considering their permeability limitations and their potential detrimental effects, such as decreased human chorionic gonadotropin secretion, elevated TNF-α levels and depressed cell viability,27 novel antioxidants have not yet been investigated. Hydrogen may have potential as a novel selective antioxidant. It has the advantages of selectively reducing toxic ROS, such as hydroxyl radical and peroxynitrite anion, and better penetration into cell membranes,28 with no detrimental effects on other free radicals and normal physiological function.29 Hydrogen may act as a therapeutic antioxidant in the prevention or treatment of PE in vivo5 and in vitro.30 At the same time, hydrogen showed no negative effects on cell proliferation and TNF-α expression,27 indicating that hydrogen is a potential antioxidant candidate in the management of PE. Our findings suggest that saturated hydrogen saline treatment effectively decreased p38 activation. We observed a significant reduction in p38 mRNA expression and p-p38 MAPK protein levels in the placentas of the hydrogen-treated groups, suggesting the involvement of MAPK cascades in saturated hydrogen saline-mediated protection. Similarly, p38 MAPK activation has been considered a potential target for therapeutic interventions to prevent adverse pregnancy outcomes mediated by stress factors,31 and a novel therapeutic target for pregnancy complications arising from placental endothelial dysfunction.32 Other drugs have been employed to suppress oxidative stress in PE by inhibiting the p38 MAPK signaling pathway, such as alpha-1-antitrypsin (AAT).33
Hydrogen, as an antioxidant agent, may affect the MAPK signaling pathway by blocking ROS, thereby reducing the inflammatory reaction and apoptosis, and ultimately improving the prognosis in HDCP. It has also been observed to improve vascular remodeling diseases via the downregulation of the p38 MAPK signaling pathway.34 In this study, hydrogen moderated degeneration and apoptosis in the placenta in the PE rat model and decreased p38 MAPK activation. Considering that placental vascular remodeling plays a critical role in PE, hydrogen may improve placental vascular remodeling by suppressing the p38 MAPK signaling pathway.
To confirm the role of the p38 MAPK signaling pathway in suppressing oxidative stress by hydrogen in PE, we employed SB203580, an inhibitor of the MAPK signaling pathway. Together with hydrogen, the LHS exhibited an even further decrease in p38 MAPK mRNA expression and p-p38 MAPK protein levels compared with the LH group. Furthermore, the LHS group showed a more pronounced depression of TNF-α and IL-1β expression compared with the LH group. By inhibiting p38 MAPK with SB203580, we demonstrated that the p38 protein plays a role in the pathology of PE. As reported, p38 inhibition by SB203580 also decreases sFlt-1/sEng secretions by suppressing oxidative stress, and activates p38 MAPK signaling.26 Activation of p38 MAPK was followed by affecting the invasion of trophoblast cells and increasing the secretions of sFlt-1/sEng, which then participate in the pathogenesis of PE.26
Limitations
First, although L-NAME, as a nitric oxide synthase inhibitor, could induce symptoms of PE in pregnant rats (including hypertension and pathological changes in the placenta and kidney), it may be difficult to accurately reproduce the pathogenesis of PE in humans. Therefore, the findings of this study should be interpreted carefully if clinical application of hydrogen in PE patients is to be considered. Second, hydrogen was administered through intraperitoneal injection in this study, but a satisfactory alternative route may need to be investigated in clinical studies.
Conclusions
Hydrogen, as an antioxidant agent, may suppress the MAPK signaling pathway by blocking ROS, thereby reducing the inflammatory reaction and apoptosis, and ultimately improving the prognosis of HDCP. Nevertheless, the potential association between the MAPK signaling pathway and oxidative stress or inflammatory responses in HDCP needs to be further clarified.
Supplementary materials
The supplementary tables are available at https://doi.org/10.5281/zenodo.7074902. The package contains the following files:
Supplementary Table 1. Shapiro–Wilk test results for protein excretion and SBP in different groups
Supplementary Table 2. Shapiro–Wilk test results for number of resorptions, fetal weight, TNF-α, IL-1β, MDA, TUNEL-positive cells, p-38 mRNA, and p-p38 protein in different groups
Supplementary Table 3. Student’s t-test for Figure 1, Figure 5, Figure 8, and Figure 9.








