
















Abstract
Background. Microbiota and tight junction proteins (TJPs) are components of the gut barrier, and are considered stress targets that have deleterious effects on intestinal homeostasis.
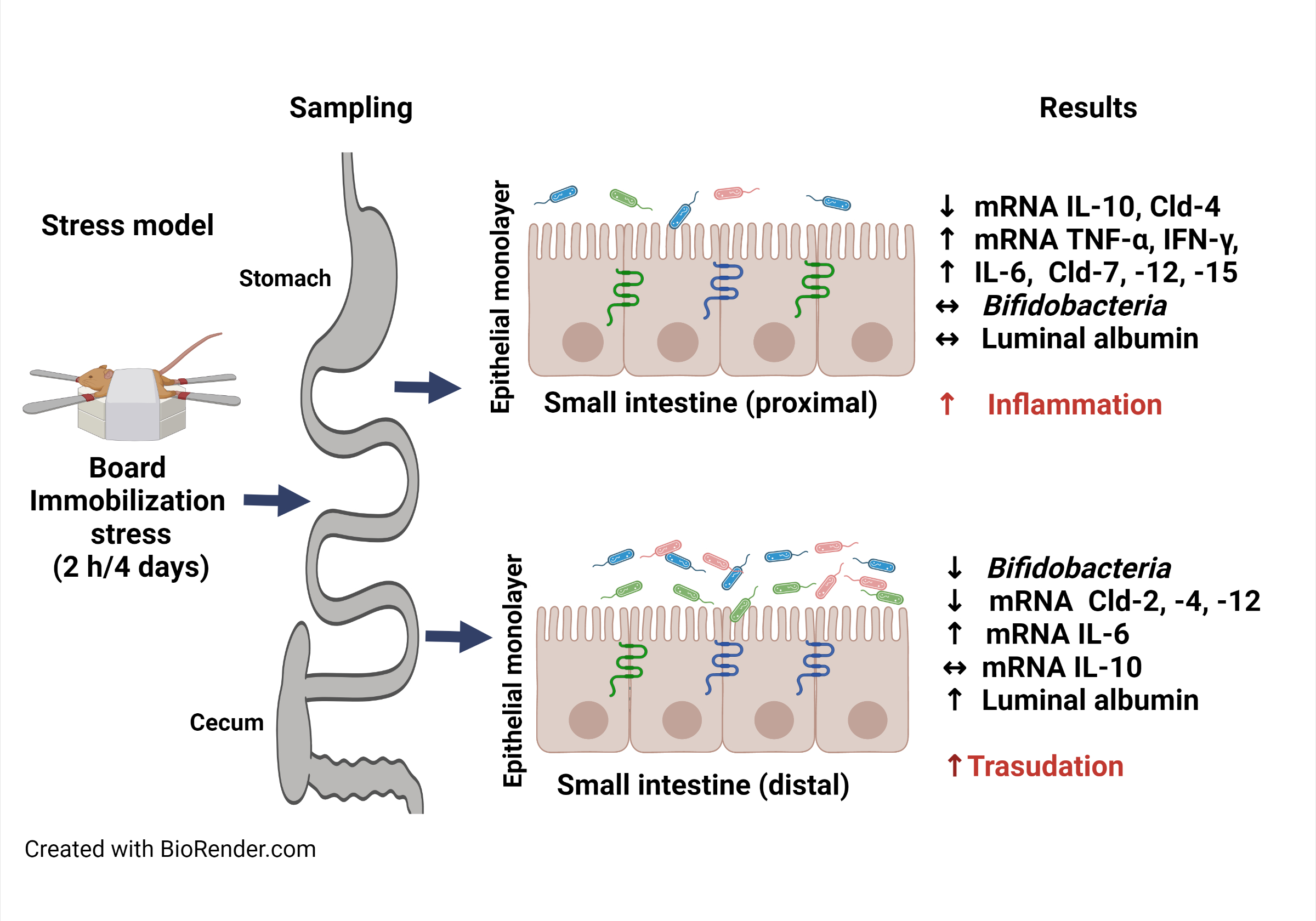
Objectives. This study aimed to evaluate the effects of chronic immobilization stress on selected small intestine homeostasis parameters.
Materials and methods. Female BALB/c mice were divided into a stress group that underwent short-term immobilization for 2 h per day for 4 consecutive days, and a non-stressed control group (n = 6 per group). Proximal and distal small intestine samples were excised to assess colony-forming units per gram (CFU/g) of total bifidobacteria in selective agar plates, luminal albumin was assessed using immune-enzymatic assay, pro-inflammatory cytokines were evaluated using reverse transcription-quantitative polymerase chain reaction (RT-qPCR), and TJPs (pore-forming, claudin (Cld)-2; pore-sealing, Cld-4; ambiguous, Cld-7, -12 and -15) were assessed with RT-qPCR and western blotting.
Results. Compared with the control group, the stress group had lower body weight and energy intake. In the distal region, the stressed mice had lower bifidobacteria count and messenger ribonucleic acid (mRNA) expression of Cld-2, Cld-4 and Cld-12, though they had more albumin and higher interleukin (IL)-6 mRNA expression. Within the proximal region, the stressed mice had higher mRNA expression of tumor necrosis factor alpha (TNF-α), interferon gamma (IFN-γ), IL-6, Cld-7, Cld-12, and Cld-15, along with lower levels of IL-10 and Cld-4. However, mRNA and protein expression of TJPs were discordant.
Conclusions. These findings indicate divergent stress-induced outcomes in the small intestine, evidenced by the elicitation of a pro-inflammatory response and decreased anti-inflammatory response in the duodenum, and by increased albumin transudation and decreased bifidobacterial growth in the distal region.
Key words: small intestine, tight junction proteins, board immobilization stress, intestinal albumin, intestinal bifidobacteria
Background
Intestinal homeostasis refers to a healthy balance of gut barrier components regarded as markers of intestinal health, including microbiota and tight junction proteins (TJPs) that bind the epithelial monolayer paracellularly.1 Gut commensals, such as those of the genus Bifidobacterium, secrete organic acids that enhance the structural and functional integrity of TJPs and prevent the deleterious effects of the potent pro-inflammatory cytokine tumor necrosis factor alpha (TNF-α) on epithelial permeability.2 Gradients in the density and diversity of such microbiota increase from the proximal to distal small intestine regions, and reach their maximum extent in the colon.3
Tight junction proteins, including occludin and claudins (Clds), are expressed at the most apical side of the epithelial cell paracellular membrane and have a regionalized location in each segment of the gastrointestinal tract,4 with occludin involved in regulating the rate of flux of molecules across the epithelial layer.5 At the same time, Clds are classified as pore-sealing, pore-forming or ambiguous, with the latter encompassing Cld-7, -12 and -13. The pore-sealing variants, Cld-1, -3, -4, -5, -8, and -18, provide high transepithelial electrical resistance (TEER), resulting in low solute permeability in the intestine. In contrast, the pore-forming proteins form channels that enable solute flow and include Cld-2 and Cld-10.6
Experimental assays demonstrate a wide array of interweaving pathways through which stress regulates commensal microbiota7 and TJPs.8 Chronic stress had a downmodulatory effect on the commensal bifidobacterial density in the colon of rats under restraint stress for 2 h a day over 4 days,9 while the abundance in metagenomic terms of Bifidobacterium pseudolongum increased in the colon of mice subjected to chronic unpredictable mild stress.10 Moreover, the interactions between chronic stress and microbiota through the brain–gut–microbiota axis impact behavior and TJP remodeling through pro-inflammatory cytokines.11, 12 Indeed, stress regulation of TJPs is partly mediated by TNF-α-induced expression of the myosin light chain kinase protein and the simultaneous opening of the gut barrier, as documented using in vitro Caco2-intestinal cultures.13
Water avoidance stress in male rats for 1 h a day over 10 days reduced the tissue location of Cld-1, occludin and zonular occludin-1 (ZO-1) in the colon but not in the jejunum. Interestingly, injecting corticosterone mimicked these effects, and they were blocked by the corticoid receptor antagonist RU-486, suggesting a regionalized stress-induced regulation of TJPs via corticosteroid receptor signaling.14 In experimental settings of chronic isolation stress in female rats, treatment with spironolactone decreased Cld-15 mRNA levels only in small intestine regardless the stress conditions indicanting a role for MR in the Cld-15 expression15 Moreover, rats subjected to subacute stress through isolation and limited movement for 24 h combined with chronic crowding stress for 14 days experienced a jejunal-specific decrease in Cld-2 messenger ribonucleic acid (mRNA) expression, with no colonic changes. However, exposure to subacute stress alone increased jejunal mRNA levels of Cld-1, Cld-5, Cld-8, occludin, and ZO-1, but did not affect the colon.16 Other studies show that chronic restraint stress for 10 h a day over 3 days in mice decreased the expression of Cld-1 and occludin in the duodenum, jejunum and ileum.17 Furthermore, chronic stress by tail suspension in rats for 14 or 21 days decreased the tissue expression and mRNA expression of occludin and ZO-1 in the jejunum.18 Previous work also described the effects of immobilization stress on TJPs in the colon19 and the contribution of stress to the progression and aggravation of pathologies affecting the small intestine.20
Objectives
This study aimed, for the first time, to assess the impact of chronic immobilization-induced stress on commensal bifidobacteria and epithelial TJPs of the duodenum and ileum by measuring pro-inflammatory and anti-inflammatory cytokines, and using albumin as a marker of permeability by assessing its extravasation from the blood to the intestinal lumen.21
Materials and methods
Animals
Twelve 6-week-old female BALB/c mice were housed at the Laboratory Animal Experimentation and Production Unit of the Metropolitan Autonomous University of Xochimilico (Mexico City, Mexico), in 2 groups of 6 mice each, under a 12-hour light/dark cycle (light on at 7:00 AM, light off at 7:00 PM), at room temperature (~20°C) and relative humidity of 55%. Mice were allowed free access to Laboratory Rodent Diet 5001 (LabDiet, St. Louis, USA) and water, and were housed for 2 weeks to adapt to the conditions. The same trained handler performed animal manipulations between 8:00 AM and 11:00 AM to reduce the influence of the circadian cycle on fluctuations in the levels of corticosterone and adrenocorticotropic hormones. The Internal Committee for the Use and Care of Laboratory Animals of the Metropolitan Autonomous University of Xochimilico approved the experimental interventions (protocol No. 176). Animals were maintained and handled following the Mexican federal regulations for animal experimentation and care (NOM-062-ZOO-1999; Ministry of Agriculture, Mexico City, Mexico), and in accordance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines.22
Body weight and energy intake
Body weight and food intake were determined on the 6th day for 7-week-old mice and on the 5th day for 8-week-old mice. The energy intake was calculated by multiplying the total food intake [g] and caloric content [kcal/g] (Laboratory Rodent Diet 5001 contains 4.675 kcal/g), and the values for each group from the 2 experiments were expressed as mean ± standard deviation (M ±SD).
Stress protocol
Eight-week-old mice were subjected to board immobilization stress for 2 h per day between the 2nd and 5th days using a previously described protocol.19 Briefly, each mouse was placed on a polystyrene board in the prone position, with all 4 limbs and the middle part of the tail gently stretched, and then attached by placing a very low-adhesive tape directly on the skin or the rear central pads, overlaid with a high-adhesive tape. Curved strips of paperboard-adhesive tape reels were placed over the tape for the mouse to chew on, preventing self-inflicted injuries of the forelimb skin. The head was allowed to move freely, and limb twisting and whisker tearing were avoided. The tape was carefully removed after 2 h of immobilization, and the mouse was released. When the experimental group underwent the stress protocol, the control mice had no access to food or water.
Sampling
Upon completion of the stress protocol, mice were euthanized via exposure to isoflurane and exsanguination by cardiac puncture. The pyloric (proximal) and cecal (distal) segments of the small intestine were dissected into 7-centimeter-long sections, and fecal content was gently removed before flushing with 1.5 mL of sterile phosphate-buffered saline (PBS) (pH 7.2), containing a cOmplete Mini™ protease inhibitor cocktail (cat. No. 11836153001; Roche Diagnostics, Mannheim, Germany). Flushing was repeated 5 times using the same liquid and the samples were collected in a Petri dish to enrich the protein content before storing the samples at −20°C for the albumin enzyme-linked immunosorbent assay (ELISA). Then, 1-centimeter intestinal segments were opened longitudinally and deposited in pre-weighed sterile microcentrifuge tubes containing 0.5 mL of thioglycolate broth (cat. No. 211651; BD Biosciences, Sparks, USA) for total count of bifidobacteria. In addition, 1-centimeter-long segments were dissected, opened longitudinally and scraped using a glass microscope slide to collect whole mucosa samples that were immediately frozen at −70°C for reverse transcription-quantitative polymerase chain reaction (RT-qPCR) and western blot assays.
Total count of bifidobacteria
Intestinal segments collected in sterile tubes containing thioglycolate broth (0.5 mL) were weighed and then fully homogenized. Intestinal homogenates were serially diluted (×10) in thioglycolate broth and 10 μL of the serial dilutions were plated on propionate agar (cat. No. 1.000430.0500) containing lithium mupirocin (cat. No. 1000.45.0010) (both from Merck, Darmstadt, Germany). After the incubation for 72 h at 37°C under anaerobic conditions using GasPack™ anaerobic sachets (cat. No. 260678) and GasPack™ dry anaerobic indicator strips (cat. No. 271051) (both from BD Biosciences), the number of colonies was counted to determine the colony-forming units per gram (CFU/g) of tissue.
Immuno-enzymatic assay
The Bradford assay quantified total protein in intestine samples using a commercial dye reagent (cat. No. 500-0006; Bio-Rad, Hercules, USA). Albumin quantitation was undertaken using sandwich ELISA in 96-well polystyrene microplates (cat. No. 8590; Costar Corning, Corning, USA) coated with goat IgG anti-mouse albumin (0.01 μg/mL) (cat. No. ab19194; Abcam, Waltham, USA) diluted in carbonate–bicarbonate buffer (pH 9.6). After the incubation overnight at 4°C, plates were washed with PBS containing 0.05% Tween 20 (PBST) (200 μL) and blocked with 5% w/v blotto (cat. No. 37530; Thermo Fisher Scientific, Waltham, USA) diluted in carbonate–bicarbonate buffer (pH 9.6), incubated for 2 h at 37°C, and then washed. The intestinal fluid samples were diluted in 5% w/v blotto in PBST, with the dilution adjusted for total protein concentration. After the incubation overnight at 4°C the wells were washed, horseradish peroxidase (HRP)-conjugated goat IgG anti-mouse albumin antibody (cat. No. ab19195; Abcam) diluted in 5% w/v blotto–PBST was added, and the plates were incubated for 1 h at 37°C. The plates were washed, a substrate solution (hydrogen peroxide and ortho-phenylenediamine in citrate phosphate buffer (pH: 5.0)) was added, and the samples were incubated for 20 min in the dark at room temperature. The enzymatic reaction was stopped with 2.5 M sulfuric acid (100 µL/well), and the absorbance was measured at 492 nm using an Epoch™ microplate reader (BioTek, Winooski, USA). An ELISA assay using purified mouse albumin (cat. No. A3139; MilliporeSigma, St. Louis, USA) was used to prepare an albumin [μg/mL] standard curve, and albumin concentration was expressed per 100 mg of total protein in the intestinal samples. All reactants and samples were tested in triplicate.
Reverse transcription-quantitative polymerase chain reaction
The RNA was isolated from proximal and distal mucosa using TRI Reagent® (cat. No. TR 118; Molecular Research Center, Inc., Cincinnati, USA), and total RNA (0.2 μg) was used as a template. The determination of mRNA expression of pro-inflammatory cytokines (TNF-α, interferon gamma (IFN-γ) and interleukin (IL)-6), an anti-inflammatory cytokine (IL-10), and TJPs (occludin, and Cld-2, -4, -7, -12, and -15) was performed using the 2−ΔΔCQ method23 described in detail by Machorro-Rojas et al.19
Western blot assay
Protein levels of Cld-2, -4, -7, -12 and -15, and occludin were assayed with western blot, according to the method described in detail by Machorro-Rojas et al.,19 using whole mucosa extracts from the proximal and distal segments of the small intestine.
Statistical analyses
All data analyses employed GraphPad Prism v. 8.0.1 software (GraphPad Software Inc., San Diego, USA), except for the energy intake and z-scores between the control and stress groups, which used a Social Science Statistics website (https://www.socscistatistics.com/tests/mannwhitney/default.aspx). Experimental assays were repeated twice, with representative data from 1 assay presented (n = 12). Data are expressed as the median with interquartile range (IQR) (Q1–Q3) (n = 6/group), and were compared using the nonparametric Mann–Whitney U test. A value of p < 0.05 indicated a statistically significant difference.
Results
Stress reduced distal bifidobacteria count, body weight and energy intake
Compared to the control group, stressed mice had a lower bifidobacteria count in the proximal segment (p = 0.522, z-score = 0.6405; Figure 1A) and a significantly lower count in the distal region (p = 0.030, z-score = 2.1617; Figure 1B). Also, body weight (p = 0.020, z-score = −2.321; Figure 1C) and energy intake (p = 0.049; Figure 1D) were significantly lower in the stress group than in the control group.
Stress displayed divergent effects on albumin and cytokine responses in each region
Albumin concentration was lower in the proximal region (p = 0.092, z-score = 1.6814; Figure 2A) and higher in the distal region (p = 0.013 z-score = −2.4819; Figure 2B) in the stress group compared to the control group. Meanwhile, relative mRNA expression of TNF-α (p = 0.008 z-score = 2.6473), IFN-γ (p = 0.005 z-score = −2.8022) and IL-6 (p = 0.008, z-score = 2.6473) were higher (Figure 2C), and the expression of IL-10 was lower (p = 0.008, z-score = −2.6473; Figure 2E) in the proximal region of the stress group. However, only mRNA expression of IL-6 significantly increased (p = 0.005, z-score = −2.8022) in the distal region of the stress group compared to the same region in the control group (Figure 2D,F).
Stress increased the mRNA expression of ambiguous claudins in the proximal small intestine
The analysis of mRNA in the proximal region (Figure 3A) demonstrated higher expression of the ambiguous Clds, Cld-7, Cld-12 and Cld-15 in the stress group than in the control group (all p = 0.005, z-score = −2.8022). Meanwhile, the mRNA expression of Cld-4 was lower (p = 0.005, z-score = 2.8022) in the stress group, and the expression of occludin (p = 0.065, z-score = −1.8414) and Cld-2 (p = 0.378, z-score = 0.8807) was nonsignificant. Western blot analysis (Figure 3B) showed increased protein levels of Cld-2 (p = 0.005, z-score = −2.8022) and lower levels of Cld-15 (p = 0.005, z-score = 2.8022) in the stress group.
Stress decreased the mRNA expression of claudins in the distal small intestine
The analysis of mRNA in the distal region of the small intestine (Figure 4A) showed lower expression of Cld-2, Cld-4 (both p = 0.005, z-score = 2.8022) and Cld-12 (p = 0.030, z-score = 2.1721) in the stress group. However, Cld-12 protein levels (Figure 4B) were lower in the stress group compared to the controls (p = 0.032, z-score = 2.1529).
Discussion
Previous research described the effects of immobilization stress on TJPs in the colon,19 so the present study focused on assessing, for the first time, the impact of chronic immobilization stress on selected markers of intestinal health and dysfunction in small intestine regions. In this regard, the study evaluated bifidobacteria, as they were found to be a reproducible and sensitive marker under the experimental conditions of stress used. Moreover, the approach is concordant with other studies in which bifidobacteria count was analyzed when monitoring the impact of stress on the intestine using conventional bacterial counts with selective agar plates.11 Under the experimental conditions of this study, stress caused a significant decrease in the amount of bifidobacteria in the distal segment of the small intestine. This finding is in line with the reduced bifidobacteria load found in the colon and feces of rats and mice that underwent chronic restraint stress.9, 11 In addition, the apparent bifidobacteria amount decrease in the proximal region may be due to the bacterial load being below the limit of detection, as this area is scarcely populated with microbiota compared to the distal segment.3
Similar to restraint and crowding stress, the stress induced in the current study promoted the loss of body weight and reduced caloric intake.11, 16, 24 Microbiota abundance is modulated by food intake and can be disrupted by increased energy expenditure under stressful conditions.7, 25 Moreover, stress-induced decreases in bifidobacteria load and body weight might contribute to anxiety and depression-like behaviors.11 Mice under chronic restraint stress had decreased acetate levels in the cecum but not in the ileum,12 and bifidobacteria are known to release acetate into the bloodstream.26 The acetate can cross the blood–brain barrier and provide neuronal inputs that impact food intake,27 while it acts as a ketogenic substrate for lipogenesis in peripheral tissues.28
The transport of mouse serum albumin from capillaries to the intestinal lumen requires crossing endothelial and epithelial barriers21 as well as selective passage through the endothelial cell membrane, followed by passive intercellular transport as serum moves into the interstitial spaces during enterocyte disintegration.29 In this study, albumin (66.5 kDa molecular mass) concentration increased in the distal region, suggesting that it is a target for stress-increased permeability. Meanwhile, chronic water avoidance stress in rats demonstrated increased polyethylene glycol-400 (400 Da molecular mass) permeability in the colon but not in the jejunum.14 Evans blue (960.83 Da molecular mass) permeability increased in the duodenum but not in the ileum or colon, of mice under acute restraint stress.30 These findings may be a result of, in part, the stress protocol used, the method of assessing permeability and the intestinal region analyzed.
The extravasation of luminal albumin was not stringently related to the mRNA expression of pro-inflammatory cytokines and TJPs in the proximal and distal small intestine regions. However, stress significantly increased mRNA levels of the mRNA of pro-inflammatory cytokines (TNF-α, IL-6 and IFN-γ) and reduced the anti-inflammatory cytokine IL-10 mRNA levels in the proximal region, whereas it only triggered increased IL-6 mRNA expression in the distal segment. Although the mRNA expression of pro-inflammatory cytokines has been shown in the colon of mice after board immobilization stress, it is unclear if these parameters have been documented in the duodenum using this model.19 Nonetheless, previous studies demonstrated increased TNF-α and IFN-γ mRNA levels and decreased IL-10 mRNA levels in the ileum after chronic restraint stress.31
Pro-inflammatory cytokines such as TNF-α, IFN-γ and IL-6, among others, contribute to enhanced gut permeability and TJP remodeling in the colon14, 32 and ileum of rats after chronic water avoidance stress33 and the ileum of mice under restraint stress.31 Moreover, pro-inflammatory cytokines play a critical role in TJP expression.34 In this study, no clear pattern was found between cytokine mRNA expression and TJPs in the proximal and distal segments. Under stress conditions, mRNA expression of Cld-4 (barrier enhancer) decreased, and of Cld-7, Cld-12 and Cld-15 (ambiguous) increased in the proximal region, whereas mRNA expression of Cld-2 (pore-forming), Cld-4 and Cld-12 was reduced in the distal region. In addition, protein levels of Cld-4 were negligible in the proximal region in both groups, while Cld-15 was detected in both regions. These findings may result from the regionalized expression of Cld-4, which is predominantly found in the distal small intestine and colon, whereas Cld-15 is distributed along the entire gastrointestinal tract.4
The findings of this study may be due to the divergent effects of stress-induced hormones on TJP expression in each small intestine region. As documented previously, TJPs in the jejunum may be regulated by transcriptional factors other than glucocorticoid receptors (GRs). Indeed, GR protein expression was lower, and TJP expression was more stable in the jejunum than colon under stress conditions, which suggests that the stress-associated decrease in TJP expression in the colon resulted from proteasome degradation.14 Moreover, a number of pathways control Cld transcription via several factors, including Snail, grainyhead-like 2 (GRHL2) and nuclear factor-kappa B (NF-κB). Furthermore, post-translational Cld modifications, including phosphorylation, ubiquitination and palmitoylation, may occur during endocytosis, exportation to subcellular compartments and proteasome degradation, among others.35
Limitations
The expression of GRs in the proximal and distal segments was not evaluated, and such data would have provided insight into the mechanisms driving the stress-induced effects.
Conclusions
The findings of this study indicate divergent outcomes of stress in the small intestine through increased pro-inflammatory and decreased anti-inflammatory cytokine mRNA expression in the duodenum. Furthermore, stress increased albumin extravasation and reduced bifidobacterial growth in the distal region of the small intestine.



