Abstract
Background. Male infertility is mostly due to low sperm quality, which accounts for about 50% of the causes of infertility. The reasons for low sperm quality are still unclear. Nowadays, many drinks contain high levels of fat, and its effect on fertility is not yet known.
Objectives. To investigate the effect of cholesterol-containing water on male fertility.
Materials and methods. Forty BALB/c male mice were divided into 2 groups: the control group and the water-induced obesity (WIO) group. Body and testicular weights were recorded and analyzed statistically. Testicular tissues were examined. Serum contents of total cholesterol (TC), triglycerides (TG), low-density lipoprotein (LDL), free testosterone (FT), luteinizing hormone (LH), and follicle-stimulating hormone (FSH) were determined. Motility count and morphology of sperm were analyzed. Real-time polymerase chain reaction (RT-PCR) was performed for SYCP3, VEGFA and WT1 genes.
Results. The results showed that the WIO group presented the highly significant values for mice body and testis weight, and TC, TG and LDL level in serum (p < 0.05), when compared to the control group. The level of FT, LH and FSH in serum was significantly decreased (p < 0.05) in the WIO group compared with the control group. Seminiferous tubules of testes became thin, and Sertoli cells showed mild atrophy in this group. Also, the count and motility of sperm significantly reduced while the ratio of sperm abnormalities significantly increased in the WIO group compared with the control group (p < 0.05). The results of RT-PCR showed that SYCP3, VEGFA and WT1 genes were significantly downregulated (p < 0.05) in the WIO group compared with the control group.
Conclusions. This study indicated that drinks containing high levels of fat may have negative effects on male fertility due to the reduction of the sexual hormones level in serum, the expression of SYCP3, VEGFA and WT1 genes, count and motility of sperm, as well as an increase in sperm abnormalities and pathological changes in the testicular tissues.
Key words: obesity, gene expression, mice, fertility
Background
The synaptonemal complex (SC) holds homologous chromosomes during the prophase of the first meiotic division1 and is essential for synapsis and meiotic recombination.2 Meiotic failure, infertility and embryonic death of mice occur due to the disruption of SC formation.3 The defective construction of SC correlates with miscarriage, infertility and Down’s syndrome in humans.4 The SYCP3 gene is the main constituent of the SC and plays a vital role in the meiosis of spermatogenesis, as well as fertility and homologous chromosome pairing in males.5 Vascular endothelial growth factor A (VEGFA) has been expressed in semen, seminal vesicles, prostate, and normal testis.6 The VEGFA is vital for blood vessel development and endothelial cell migration. In addition, it regulates the spermatogonial stem cell pool and male reproductive lifespan.7 Wilms’ tumor gene (WT1) has been expressed in testis, kidneys and ovaries.8 It has an important role in ovarian follicle development9 and spermatogenesis.10
Previous studies indicated that obesity correlated with infertility; it has been observed that bodyweight is significantly increased in patients with male factor infertility.11 In recent years, overweight in adults males was correlated with low semen quality.12, 13 Obesity has been associated with subfertility and was shown to be related to long waiting time to pregnancy.14 Over the past half-century, several scientific reports had proven that the decrease in semen quality and male reproductive capacity has occurred in parallel with the increase in obesity rates,15 indicating the need to focus on the probability of obesity as a cause of male infertility and reduction in fecundity. Testosterone deficiency may contribute to a decrease in semen quality and total sperm count in obese men.16 The previous epidemiological studies suggested a significant negative association between high body mass index (BMI) and the semen parameters, involving semen volume,17 and sperm concentration,18 motility19 and morphology.20 Spermatogenesis is affected by altered levels of sex hormones in obese men, such as decreased free or total testosterone levels and increased estradiol levels in serum.21 Moreover, diet-induced obesity increases the DNA fragmentation index in spermatozoa, causing clear weakening of male fertility.22
The previous studies demonstrated the negative effect of high-fat-diet-induced obesity on fertility, but the mechanism of how obesity can cause male subfertility was poorly characterized. In this study, we initially established water-induced obesity (WIO) model in order to determine whether obesity affects the decline of male fertility as well as serum reproductive hormone levels, sperm morphology, and/or disrupts testicular morphology and expression of some genes related to male fertility.
Objectives
Although changes in the sexual hormones and the count, motility and quality of sperm, as well as testicular histopathology accompanied by increasing adiposity have been identified by several previous studies on high-fat diet, the effect of WIO on the sexual hormones level and count, motility and morphology of sperm, testicular histopathology and gene expression related fertility remain unknown. Also, to our knowledge, this is the first study focusing on the effect of obesity on the expression of SYCP3, VEGFA and WT1 genes in testis.
Materials and methods
WIO model
To study the effect of obesity on the expression of some genes related to fertility, BALB/c mice model was developed, in which obesity was enhanced by adding 0.5 g of cholesterol and 0.1 g of ox gall to 400 mL of drinking water manipulation, in order to model the human condition. After BALB/c mice were given drinking water containing a high level of cholesterol for 70 days, the WIO model was successfully established.
Experimental design
The experimental procedure used in this investigation was approved by the Animal Care and Use Committee of National Research Centre (Cairo, Egypt; approval No. NRCE-CBD1992020). Forty BALB/c male mice aged 2 weeks, weighing 21−25 g, were divided into 2 groups; each group included 20 mice housed in cages maintained at 22 ±2°C with 50 ±5% humidity and 12 h light/dark cycle. The control group was fed normal diet for 70 days; the WIO group was fed normal diet and 0.5 g of cholesterol with 0.1 g of ox gall in 400 mL of drinking water for 70 days. During this period, individual body weight for the 2 groups was measured every 2 weeks. At the end of experiment, all mice were sacrificed by neck vertebra luxation. The testes of each mouse were taken and weighted; one testis was stored in 10% formalin for the histopathological examination, and the second testis was stored in −80°C for the examination of gene expression.
Measurement of lipid profile
and sexual hormones
Blood samples were collected from the eyes of all mice from the control group and the WIO group at the end of the experiment, and then centrifuged at 5000 rpm for 10 min to collect serum. A specific kit (MAK045, RAB0734) bought from Sigma-Aldrich (St. Louis, USA) was used to measure total cholesterol (TC), triglycerides (TG) and low-density lipoprotein (LDL) levels at 546 nm with a spectrophotometer (UV-240; Shimadzu, Kyoto, Japan). Free testosterone (FT), luteinizing hormone (LH) and follicle-stimulating hormone (FSH) levels were determined using competitive immunoassay technique.23
Testicular histopathology
Testicular tissue samples were taken from all mice from the control group and the WIO group. The samples were fixed in 10% formalin, processed and stained with hematoxylin and eosin (H&E). Histopathological studies using light microscopy and photomicrographs were made.
Sperm parameter
At the end of the experiment and after sacrificing the mice, the epididymides were removed from each mouse. Then, sperm was collected in Petri dish containing 1 mL of phosphate buffered saline at 37°C through cutting cauda of epididymides. Motility and count of spermatozoa were calculated using hemocytometer and a drop of a homogenate smeared on a cleaned slide. These slides were left to dry and coded, and then stained with approx. 0.05% aqueous eosin Y solution. About 1000 sperm cells were examined to detect morphological abnormalities in sperm head and tail for each mouse.
Gene expression
Testicular tissues were homogenized using a homogenizer (Z722375, IKA 3703100; Thermo Fisher Scientific), and suspended in TRIzol® (Thermo Fisher Scientific) in order to extract total RNA for quantitative real-time polymerase chain reaction (qRT-PCR). The NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, USA) was used to measure the quantity and quality of RNA. The cDNA synthesis was performed using COSMO cDNA synthesis kit (WF10205002; Willowfort, Birmingham, UK), according to the manufacturer’s instruction. Real-time polymerase chain reaction was performed on 4 host genes (SYCP3, VEGFA, WT1, and β-actin). The primers were designed using National Center for Biotechnology Information (NCBI) primer BLAST, and checked using Oligo v. 7 (Molecular Biology Insights, Inc. (DBA Oligo, Inc.), Colorado Springs, USA). All primers were synthesized by Macrogen, Inc. (Seoul, South Korea) (Table 1). The gene expression was normalized to mice β-actin and analyzed using EvaGreen® qPCR Mix Plus (ROX) (Solis BioDyne, Tartu, Estonia) using Stratagene Mx3000P RT-PCR system (Agilent Technologies, Santa Clara, USA).
Statistical analyses
There are 3 different types of data obtained from the experiment. Non-normal data distribution (sperm morphology) was analyzed using Mann–Whitney test. Normal data distribution had unequal variances between the groups (FT and FSH analysis, and the 1st week, 2nd week, 6th week and 8th week body weight measurement) and was analyzed using Welch’s test. Normal data distribution had equal variances in the group (lipid profile, LH and the other measurements of body weight) and was analyzed using independent t-test. The normality distribution was tested using Shapiro–Wilk test (SPSS v. 18 software (SPSS Inc., Chicago, USA)). The analyses of the relative quantification by RT-PCR were performed using the 2–ΔΔCT value method. The value of p < 0.05 was considered statistically significant.
Results
Body and testicular weight
Forty mice were included in the experiment, divided into a control group (n = 20, weight: mean ± standard deviation (SD) = 23.3 ±0.51 g) and a WIO group (n = 20, 23.2 ±0.5 g), and they did not present any significant differences in body weight between the 2 groups. During the first 7 days, mice body weight non-significantly increased in the WIO group (25 ±0.3 g) compared with the control group (24.6 ±0.5 g). The difference in body weight of mice between the WIO and control became significant after 2 weeks (14 days) of drinking water provided to them, and persisted for the subsequent 70 days (p < 0.001). Also, testicular weight significantly increased in the WIO group (n = 20, 0.12 ±0.02) compared with the control group (n = 20, 0.1 ±0.003) (Table 2).
Lipid profile and sexual hormones
Five replicates of serum samples were used to determine TC, TG, LDL, FT, FSH, and LH levels in each group, and each replicate was collected from 4 mice. Total cholesterol (230.2 ±5.8 mg/dL), TG (210.4 ±5.6 mg/dL) and LDL (135.8 ±2.2 mg/dL) significantly increased in the WIO group compared with the control group (125.2 ±3.9 mg/dL, 98.8 ±4.2 mg/dL and 50 ±2.7 mg/dL, respectively). Free testosterone (0.1 ±0.01 pg/dL), FSH (0.04 ±0.03 pg/dL) and LH (0.02 ±0.01 pg/dL) significantly decreased in the WIO group compared with the control group (30.8 ±1.3 pg/dL, 1 ±0.16 pg/dL, and 0.02 ±0.03 pg/dL, respectively) (Table 3).
Sperm motility and count
Sperm motility was reduced by 50% in the WIO group compared with the control group. Sperm count of obese mice was recorded as 16 × 106/mL, while for the control group it was 40 × 106/mL.
Sperm morphology
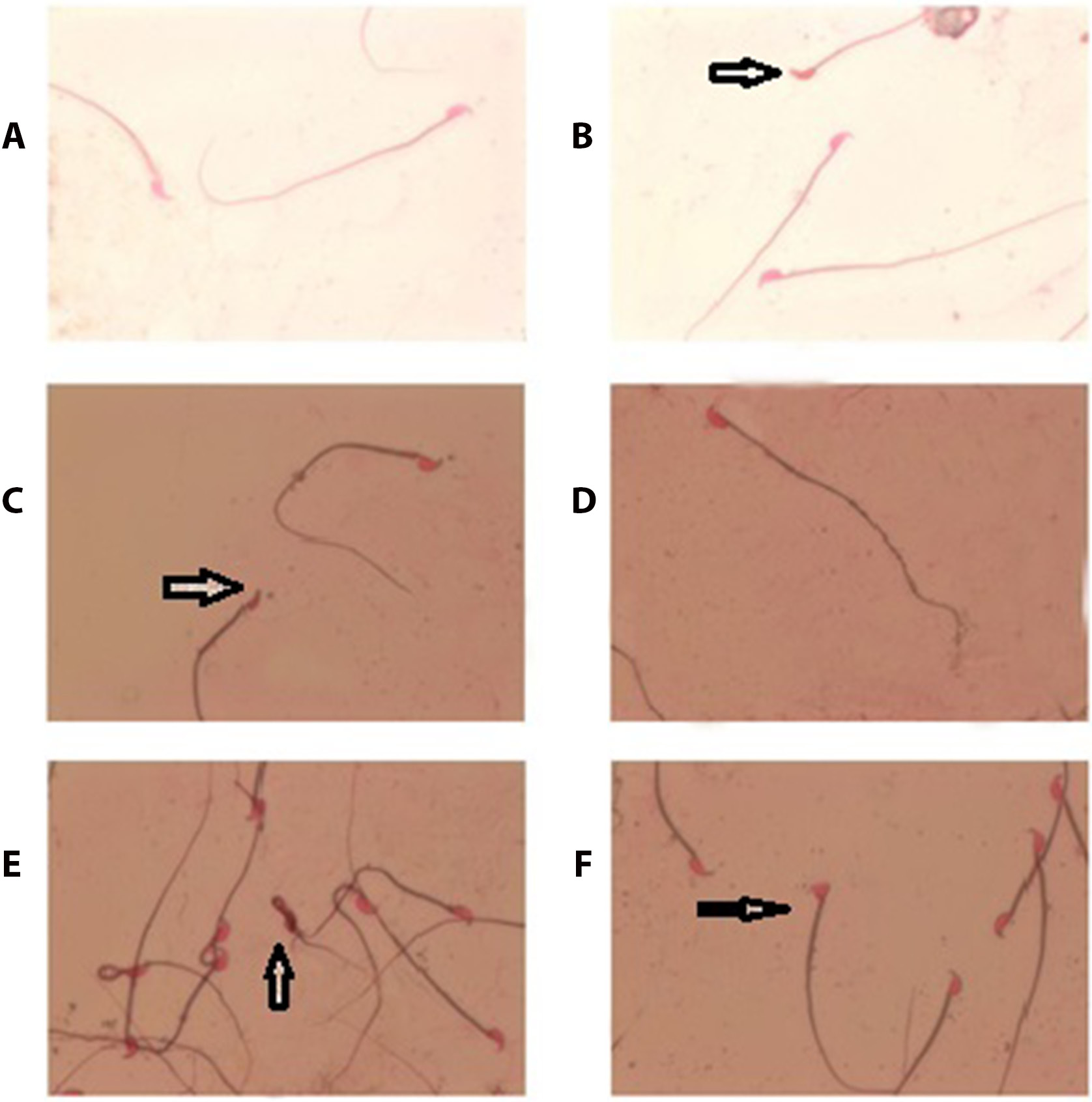
Sperm morphology was normal by 83.9% and abnormal by 16.1% in control group, while WIO group presented 58.6% normal morphology and 41.4% abnormal morphology. Median with 1st and 3rd quartile of normal sperm in WIO group was (585.5; 584,587.5), without hock abnormality (24.5; 24,25), small head (148; 146,150), amorphous head (76; 75,77), banana head (16; 15,17.75), triangle head (10; 9,12) and coiled tail (138; 137,140), compared with control group ((839; 834.4,844.1), (12.2; 11.3,13), (48; 46,50), (32; 31,32.6), (4; 3,5), (2; 1,3.75), and (62; 61,63), respectively)) (Table 4 and Figure 1).
Testicular histopathology
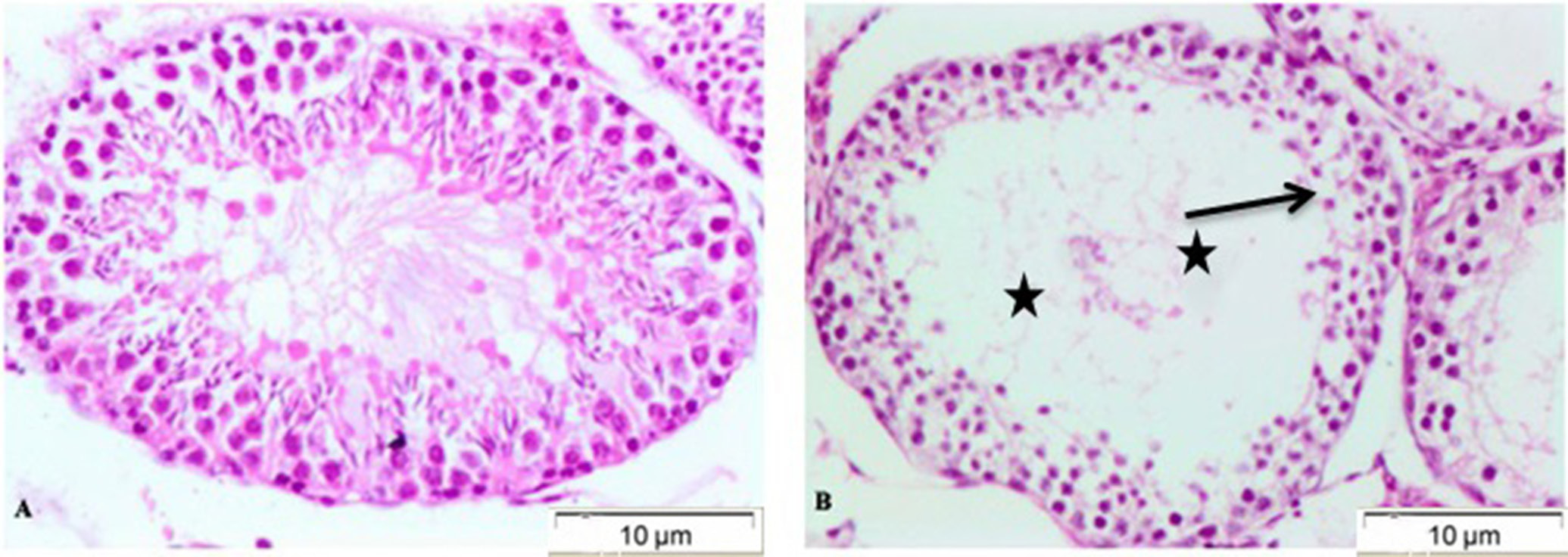
Testicular tissue of all mice was stained with H&E to confirm the effects of WIO on morphological changes in this tissue. The analysis of images obtained with light microscopy showed some changes that occurred in the testicular cells. The structure of the seminiferous tubules was normal and complete, with slight edema in the Leydig cells of the control group (Figure 2A), while in the WIO group, seminiferous tubules showed mild to severe depletion of spermatozoa, spermatogonial cells showed severe sloughing, and mild atrophy of Sertoli cells has been noticed (Figure 2B).
Gene expression
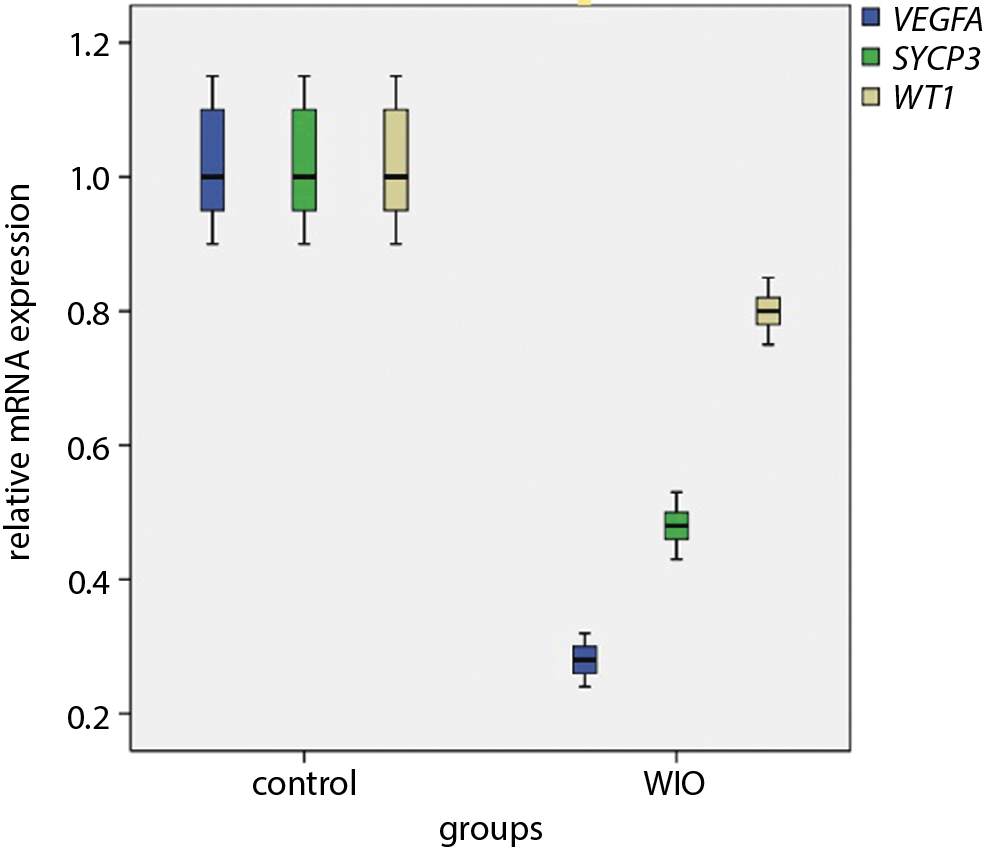
Expressions of VEGFA gene (0.28) and SYCP3 gene (0.48) were highly significantly downregulated in the WIO group compared with the control group. At the same time, the expression of WT1 gene (0.8) was significantly downregulated in the WIO group compared with the control group (Figure 3).
Discussion
Infertility is one of the causes of psychological problems for people and production deficiency in farm animals. Based on epidemiological data, an average of 10% of the world’s population of the reproductive age suffers from infertility.24 Fifty percent of cases of male infertility occur due to the low quality of sperm, which has become a source of concern in the world.25 Several studies reported the association between obesity and male infertility.11 The high-fat diet may have a critical role in metabolic diseases, causing an increase in TC, LDL and high-density lipoprotein (HDL) levels, accompanied by a significant body weight gain. In a study by Fan et al., the increase of body weight in mice fed on a high-fat diet became significant after 3 weeks.26 Dardmeh et al.27 found that an increase of body weight of mice fed a high-fat diet became significant after 4 weeks. Moreover, testicular weight significantly increased in these mice compared with the control group. In the present study, the body weight of the WIO group mice significantly increased compared with the control group after 2 weeks. Also, the testicular weight showed a significant increase in the WIO group. The serum contents of TC, TG and LDL in the WIO group increased significantly in comparison with the control group. Total cholesterol, TG and LDL significantly increased in mice fed a high-fat diet.28 In obese and hyperlipidemic mice, the level of testosterone hormone in serum was significantly decreased, and the same results were shown in humans.29 In the present study, the TH level in serum significantly decreased in the WIO group compared with the control group. This is probably due to the harmful and degenerative effects of high cholesterol levels on the secretory ability of Leydig and Sertoli cells.30 Also, our results showed a significant reduction of LH and FSH in WIO group compared with the control group, in agreement with the previous studies on diet-induced obese (DIO) mice model.27 The decrease in LH and FSH levels may be correlated with the alteration of androgens to estradiol, resulting in increased serum estrogen levels.31 The higher levels of estradiol in DIO models decrease the production and secretion of LH and FSH, causing a reduced testicular function and the testosterone production.32 Also, it was suggested that exuberant estradiol has direct harmful effect on spermatogenesis.32 In this study, we found that count and motile of sperm significantly decreased in the WIO group compared with the control group. In addition, the presence of abnormalities in shape of sperm head and tail in WIO group was significantly higher than in the control group, in which a significantly higher percentage of immotile sperm was recorded,33 altering the spermatogenesis process and affecting sperm maturation.31 The percentage of progressively and nonprogressively motile sperm significantly decreased in the high-fat diet group, which recorded the significantly higher percentage of immotile sperm.34 Although many studies have been conducted to investigate the negative effect of obesity on fertility, sperm quality, quantity and motility, as well as its effect on sexual hormones, there is not a single study on the effect of obesity on the expression of genes associated with fertility. The SYCP3 plays an important role in meiosis of spermatogenesis and fertility.5 Mice deficient in SYCP3 fail to establish synapses, leading to meiotic disruption during male spermatogenesis.35 The VEGFA regulates spermatogonial stem cell pool and male reproductive lifespan.7 The VEGFA is the first selective tissue-angiogenic molecule that stimulates the existence and proliferation of the endothelial cells of testis, and may aid in the entry of hormones into the vascular system.36 The WT1 gene is associated with ovarian follicle development9 and spermatogenesis.10 In the present study, the expressions of VEGFA, SYCP3 and WT1 genes were significantly decreased in the WIO group compared with the control group. In addition, histological analysis of obese mice testis showed the leanness of the seminiferous tubules, a clear deterioration of sperm cells within the seminiferous tubules with the formation of seminal giant cells, interstitial edema, and necrosis of Leydig cells. The VEGFA isoforms produced by Sertoli and germ cells are needed to maintain sperm quality and normal fertility.37 These results may help in explaining the relationship between the pathological changes in testicular tissues of obese mice and the downregulation of VEGFA, SYCP3 and WT1 genes.
Limitations
In this study, we encountered 1 limitation. The amount of blood taken from each mouse was small, and therefore the volume of serum extracted from it would also be very small and insufficient for all biochemical analyses. Thus, the solution was to collect 4 blood samples from each group into 1 tube, providing 5 replicates for each group. Although we could not obtain results of all biochemical analyses from each animal individually, the outcome of our study is consistent with the previous studies about the effect of obesity on the level of the sexual hormones and TC, TG, LDL, and HDL in serum.
Conclusions
Water-induced obesity led to an increase in body and testicular weight, and in the serum content of TC, TG, and LDL. In addition, it reduced the level of FT, LH and FSH in serum. Seminiferous tubules of obese mice testis became thin and showed mild to severe depletion of spermatozoa. Also, Sertoli cells showed mild atrophy. The WIO caused the downregulation in the expression of SYCP3, VEGFA and WT1 genes associated with spermatogenesis, the abnormality in the sperm shape of either head or tail, and reduced count and motility of sperm. Therefore, this study recommends limiting the amount of drinks containing high percentage of fat.