Abstract
Background. SIRT1 plays a protective role against diabetic retinopathy as it regulates inflammation, apoptosis and autophagy of cells.
Objectives. This study was designed to investigate the effects of arbutin and to identify a potential mechanism of action. Adult human retinal pigment epithelial (ARPE-19) cells were exposed to high glucose (HG) or treated with different concentrations of arbutin.
Materials and methods. The protein levels of pro-inflammatory cytokines, like tumor necrosis factor-α (TNF-α), interleukin (IL)-1β), IL-6, and p65 were assessed using enzyme-linked immunosorbent assay (ELISA). The expression of NF-κB p65 and cyclooxygenase-2 (COX-2) was detected with western blot assay. Cell apoptosis was analyzed with TUNEL assay, and expression levels of Bcl2, BAX, cleaved caspase-3, cleaved PARP, LC3II, LC3I, and beclin1 were detected with western blot assay. Autophagy levels were detected using LC3II immunofluorescence staining.
Results. Arbutin treatment markedly enhanced viability and autophagy mediators, decreased pro-inflammatory proteins and reduced apoptosis in ARPE cells under HG exposure, while increasing SIRT1 protein level. This could be blocked by Sirtinol treatment. Additionally, 3MA treatment significantly reduced the efficacy of arbutin against inflammatory markers and apoptosis in ARPE cells exposed to HG.
Conclusions. Arbutin suppressed inflammation and apoptosis of ARPE cells induced by HG by promoting autophagy via SIRT1. A potential target, SIRT1, was identified for the treatment of DR, and new effects of and action mechanisms for arbutin were found and confirmed.
Key words: arbutin, SIRT1, p65, autophagy, diabetic retinopathy
Background
Diabetic retinopathy (DR) is a serious complication of diabetes.1, 2 It begins with macular edema, which ruptures the blood–retina barrier and eventually leads to the formation of a hard exudate. Chronic hyperglycemia is the basis of DR pathogenesis.3 Retinal pigment epithelium, located between the neuroretina and choroid, is an important dense pigment layer in the retina. It plays an important role in maintaining the structural and functional integrity of the retina,4 as well as a crucial role in the pathogenesis of DR.5, 6
Arbutin is a natural soluble glycosylated phenol with the molecular structure C12H16O. It exhibits a wide range of pharmacological activities, including anti-inflammatory and antioxidant effects.7, 8, 9, 10 Studies have shown that arbutin can protect HK-2 cells from damage induced by high glucose (HG),11 and reduce H2O2-induced damage to optic ganglion cells.12 However, whether arbutin can inhibit retinal damage induced by HG remains to be investigated. Studies have shown that arbutin can inhibit the effect of lipopolysaccharide-induced lung injury by activating silent mating type information regulator 2 homolog 1 (SIRT1).13 The SIRT1, which is a NAD+-dependent class III histone deacetylase, plays a wide range of roles in retinal injury induced by HG, which could mediate the regulation of NF-κB signaling. It has been shown to regulate the cell apoptosis pathway in HG-treated cells.14, 15, 16, 17, 18, 19, 20
Objectives
The purpose of this study was to explore the mechanism of arbutin in reducing inflammation and apoptosis of retinal cells induced by HG.
Materials and methods
Cell line
Adult human retinal pigment epithelial (ARPE-19) were purchased from American Type Culture Collection (ATCC, Manassas, USA). The ARPE-19 cells were cultured in DMEM/F12 medium (Hyclone, Logan, USA) containing 10% fetal bovine serum (FBS; Gibco, Waltham, USA), in a cell incubator at a constant temperature of 37°C, containing 5% CO2. The cell status was observed daily, and the medium was changed every 2 days.
MTT assay
The ARPE-19 cells at logarithmic growth phase were seeded into 96-well plates with a cell density of 1 × 104/mL (200 μL of cell suspension; Corning Inc., Corning, USA). After the cells fully adhered to the well, the culture medium was removed and replaced with a high-sugar treatment; for the control group, this comprised normal culture medium, while the rest of the treatment groups, medium was used with a glucose concentration of 50 mmol/L. Cells were cultured for 24 h or 48 h. The medium was removed and MTT was added. Cells were cultured at 37°C for 4 h, then dimethyl sulfoxide (DMSO) was added to fully dissolve the crystals. The optical density (OD) at 490 nm was detected using a microplate reader (SpectraMax M5; Thermo Fisher Scientific, Waltham, USA).
ELISA
The ARPE-19 cells (5 × 104/mL) at logarithmic growth phase were seeded into cell culture flask. Twenty-four hours after adherent fusion, the cells were treated with different concentrations of arbutin for 24 h or 48 h. The culture medium was collected and centrifuged at 3000 rpm for 10 min to obtain the supernatant. The protein levels of pro-inflammatory cytokines, like TNF-a, IL-1β, IL-6, and p65 were assessed using ELISA assay. The expression of NF-κB p65 and cyclooxygenase-2 (COX-2) was detected with western blot assay. The absorbance at 450 nm was detected using a microplate reader (SpectraMax M5).
Western blot
The ARPE-19 cells in each group were collected and digested using RIPA Lysis Buffer containing phenylmethylsulfonyl fluoride (PMSF; 80 μL) for 30 min. The supernatant was collected in a high-speed bench refrigerated centrifuge. The concentration of protein was detected using the BCA method. After SDS-PAGE electrophoresis, the protein was transferred to a polyvinylidene difluoride (PVDF) membrane, which was blocked using bovine serum albumin (BSA) for 1 h and then washed with Tris-buffered saline with Tween (TBST) 3 times. The primary antibody (NF-κB p65, ab32536, 1 : 30; COX-2, ab169782, 1 : 2000; Bcl-2, ab32124, 1 : 1000; Bax, ab182733, 1 : 2000; cleaved caspase-3, ab32042, 1 : 500; GADPH, ab8245, 1 : 2000; all from Abcam, Cambridge, UK; and cleaved PARP, #9541, 1 : 1000; Cell Signaling Technology, Inc., Danvers, USA) was produced using the corresponding dilutions detailed above, added to the membrane and incubated at 4°C overnight. The membrane was then washed with TBST 3 times on the 2nd day. The secondary antibody conjugated with horseradish peroxidase (HRP; ab7090, ab97040, 1 : 10000; Abcam) was then added and incubated with the membrane in a shaker at room temperature for 1 h. GAPDH was used as an internal reference.
TUNEL assay
Apoptotic cells were stained with FITC-labeled dUTP using terminal deoxynucleotidyl transferase as a catalyst. The experiment was performed using a TUNEL kit (Roche, Basel, Switzerland) according to the manufacturer’s protocol. DAPI was used to stain the nucleus, and results were observed under a fluorescence microscope (Olympus Corp., Tokyo, Japan).
Immunofluorescence assay
The cells were fixed with 100% methanol for 5 min at room temperature and then permeabilized with Triton X-100 (3 g/L) for 30 min. The primary antibody (anti- LC3II; Abcam) was used with cells overnight at 4°C. Next, the secondary antibody (an Alexa Fluor® 488 goat anti-rabbit secondary antibody; Abcam) was added for incubation for 30 min at room temperature. DAPI was used to stain the nucleus. Anti-fluorescent quenching agent was added to mount the sections. The cells were observed under a fluorescence microscope (Olympus Corp.).
Statistical analyses
Summary results were expressed as mean ± standard deviation (SD). Each experiment was repeated 3 times or more within each group. Data comparing more than 2 groups were analyzed with one-way analysis of variance (ANOVA), followed by Tukey’s test. Each experiment was repeated at least 3 times. A value of p < 0.05 indicated that differences between groups were statistically significant. Statistical analysis was performed using Graph Pad Prism v. 8.0 software (GraphPad Software, San Diego, USA).
Results
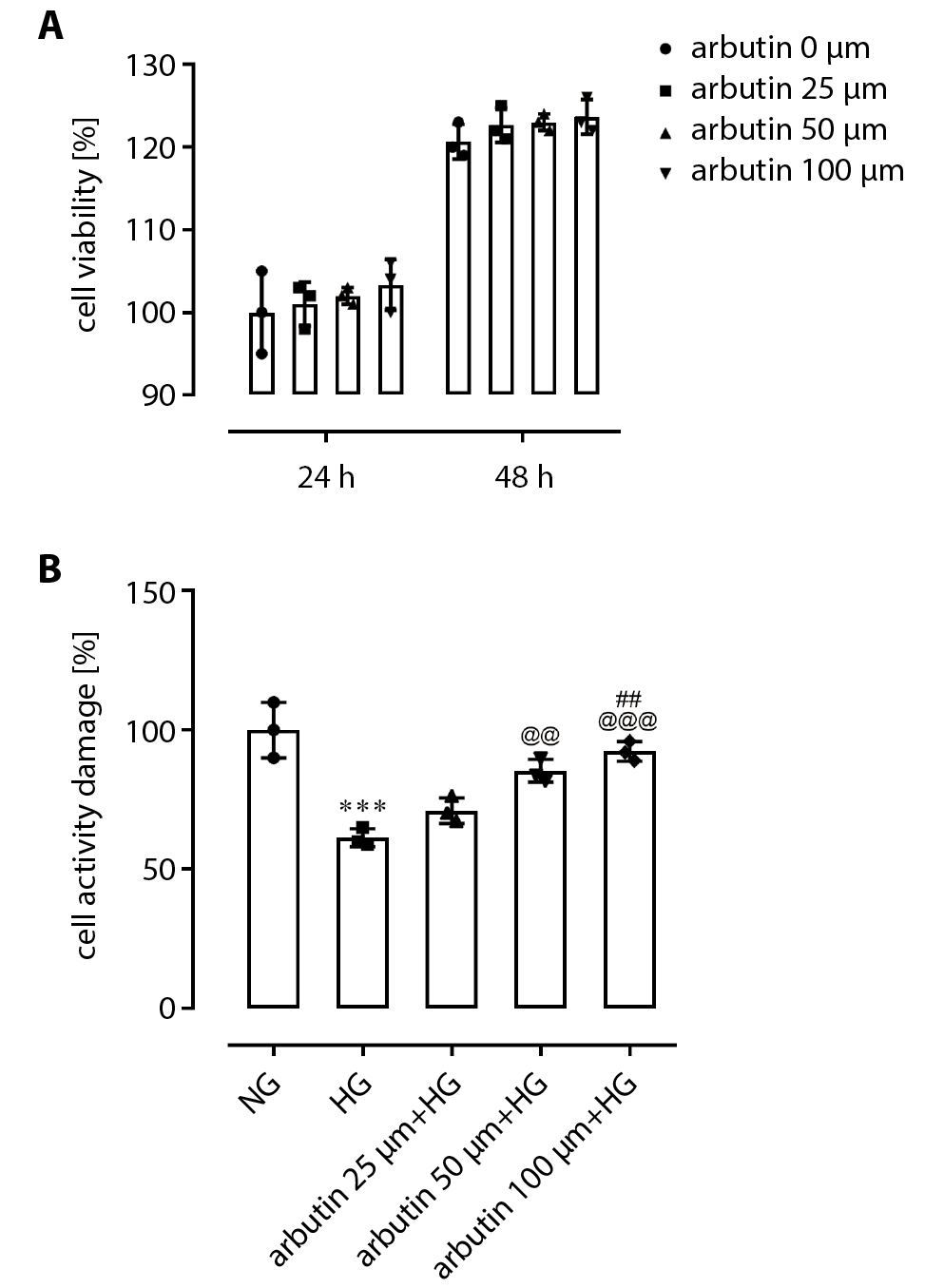
The effects of arbutin on APRE19 cell viability following HG exposure
Retinal pigment epithelial cells are regarded as the most important cells implicated in DR. A MTT assay was used to detect the effects of different concentrations of arbutin (25 μm, 50 μm or 100 μm) on the cell viability of APRE19 cells. Results showed that there was no statistically significant difference in the viability of APRE19 cells treated with 25 μm, 50 μm or 100 μm arbutin after 24 h or 48 h compared with the control group (0 μm) (Figure 1A). When the cells were pretreated with 25 μm, 50 μm or 100 μm arbutin for 1 h and then treated with HG (30 mM), the activity of ARPE-19 cells was significantly higher than that of the HG group in a dose-dependent manner (Figure 1B).
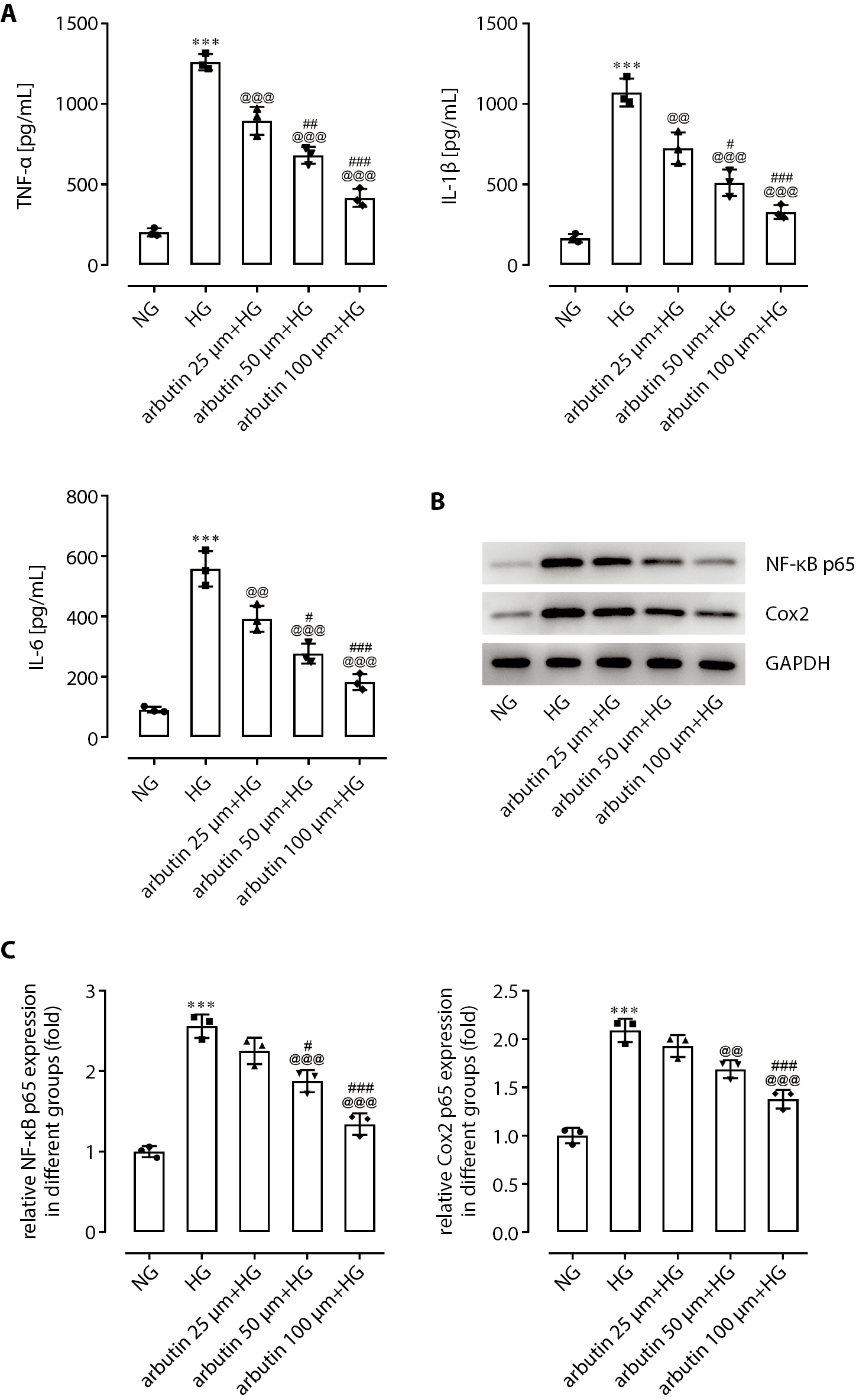
The effects of arbutin on the NF-κB pathway
As shown in Figure 2A, the expression levels of TNF-a, IL-1β and IL-6 in the APRE19 cell supernatant treated with 30 mmol/L HG for 24 h were significantly higher than those of the control group. After pretreatment with arbutin, APRE19 cells were cultured in a high-sugar environment, and TNF-α, IL-1β, and IL-6 levels in the cell supernatant were significantly reduced in a dose-dependent manner. The protein expression levels of NF-κB p65 and COX-2 in APRE19 cells were assessed using western blotting assay. The expression levels of NF-κB p65 and COX-2 were significantly increased by different modes (25 μm, 50 μm or 100 μm) of arbutin treatment (Figure 2B,C).
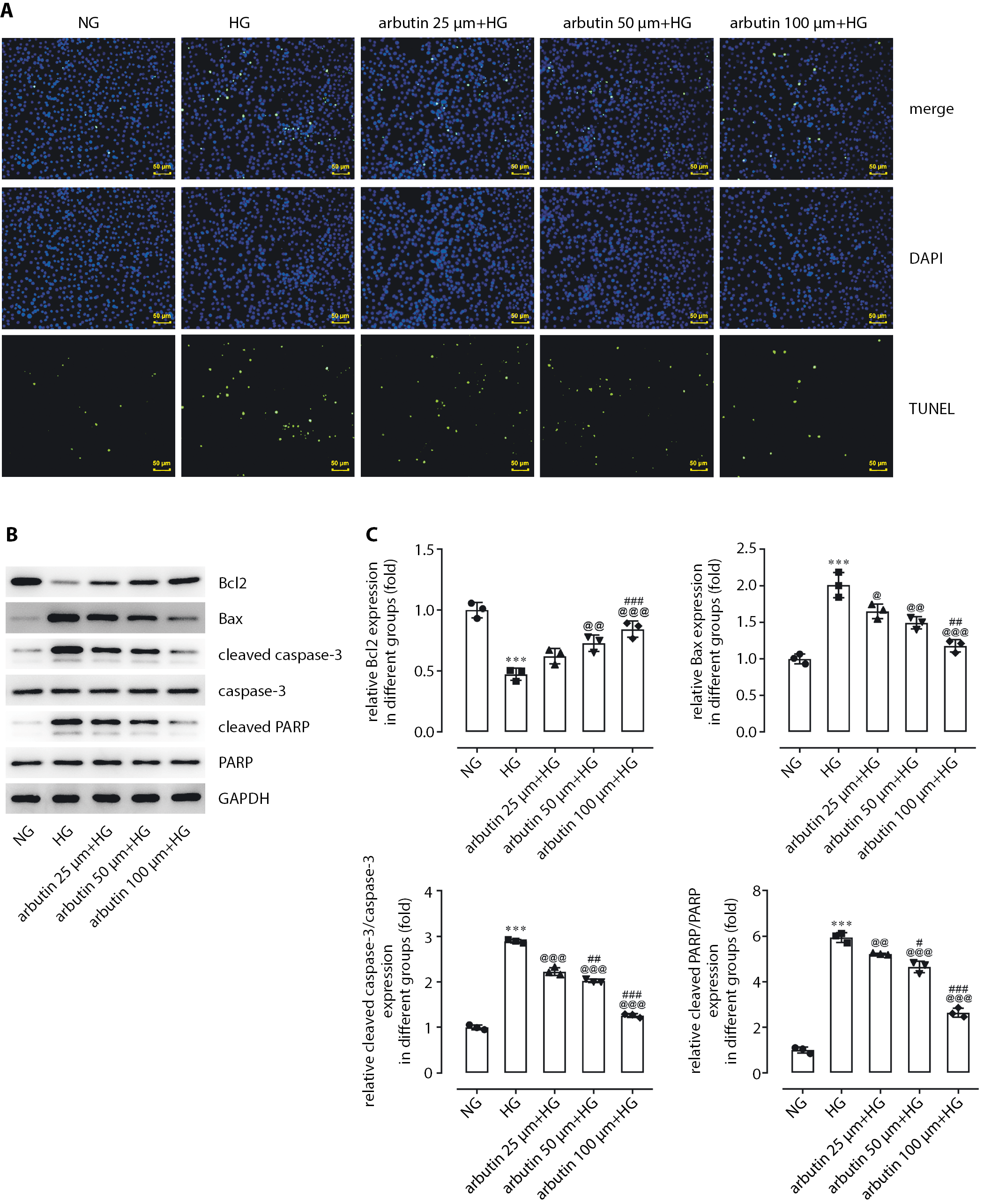
Arbutin inhibition of apoptosis
in APRE19 cells under HG exposure
A TUNEL assay was used to detect the effects of different concentrations of arbutin (25 μm, 50 μm or 100 μm) on apoptosis in APRE19 cells. Compared with the HG group, APRE19 cells treated with arbutin showed gradually decreasing apoptosis levels with increasing arbutin concentration (Figure 3A). To identify the potential mechanism by which arbutin suppressed cell apoptosis, levels of Bcl2, BAX, cleaved caspase-3 and cleaved PARP were detected through western blotting assay (Figure 3B,C). The results show that arbutin led to a significant reduction in the expression of BAX, cleaved caspase-3 and cleaved PARP in APRE19 cells compared with the HG group. On the contrary, the expression of Bcl2 was increased.
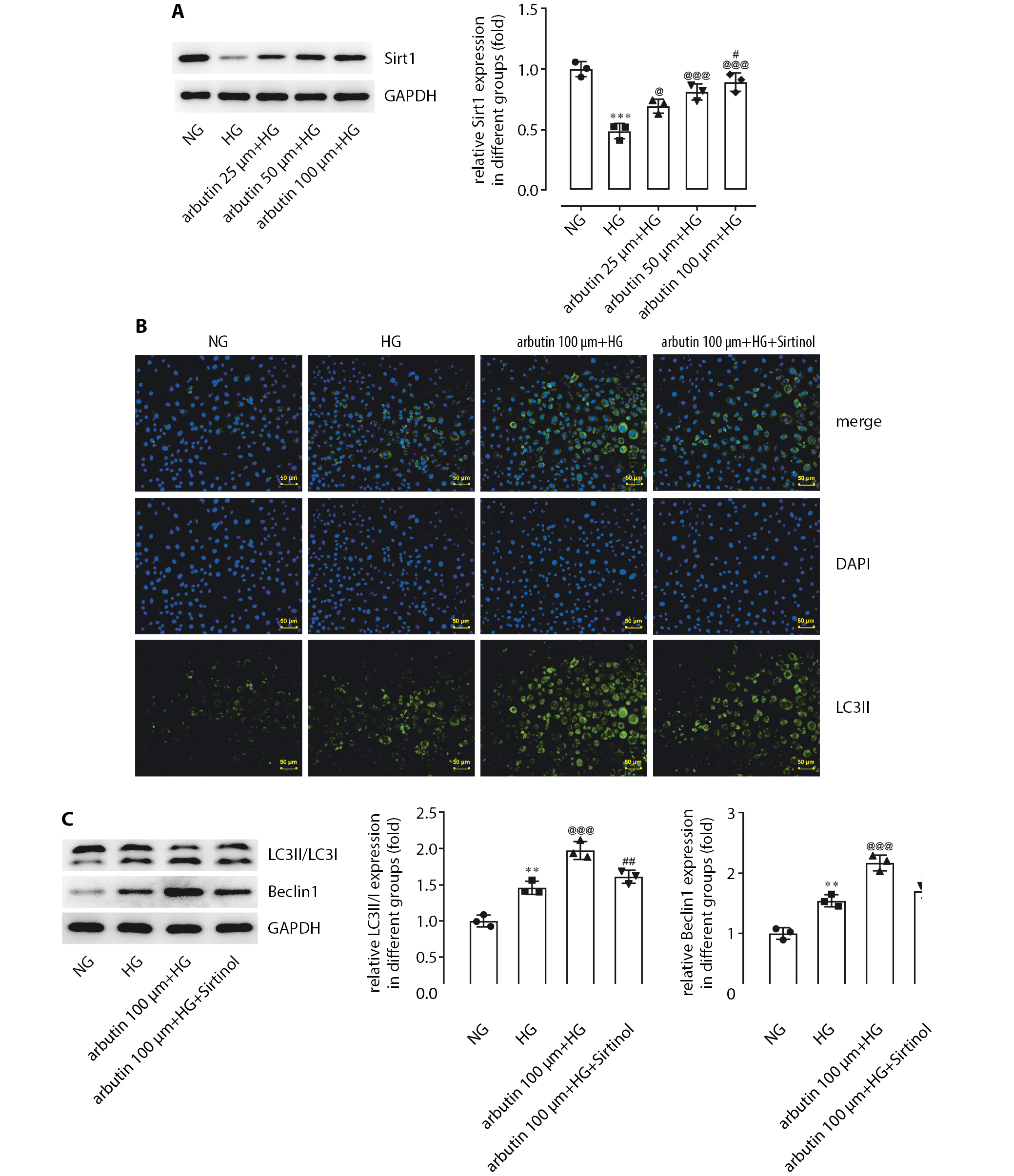
SIRT1 inhibition interferes
with arbutin suppression of autophagy
The above results fully demonstrated that arbutin could suppress both the activity and levels of inflammatory mediators, and cell apoptosis caused by HG. However, the exact mechanism of the action of arbutin was unknown. A study has suggested that SIRT1 can reduce inflammation and apoptosis of cardiomyocytes by promoting autophagy.21 Therefore, we investigated whether SIRT1 could mediate the effects of arbutin on the inflammation and apoptosis of APRE19 cells. Western blotting was used to detect the changes of SIRT1 protein expression in APRE19 cells after arbutin treatment at different concentrations (25 μm, 50 μm or 100 μm). The HG exposure significantly reduced the relative protein level of SIRT1 in APRE19 cells when compared to the normal glucose (NG) group (Figure 4A). Additionally, compared to the HG group, the expression level of SIRT1 in APRE19 cells significantly increased in response to increased arbutin concentration (Figure 4A).
Sirtinol (100 μm), an inhibitor of SIRT1, was used to pretreat APRE19 cells for 1 h, after which APRE19 cells underwent a combination treatment of HG and arbutin (100 μm) for 24 h. The immunofluorescence results showed that expression of LC3ll in the HG group was higher than expression in the normal group (Figure 4B). LC3ll expression was further increased by co-treatment with arbutin and HG. Surprisingly, sirtinol significantly suppressed the effects of arbutin.
Western blot results showed that the expression level of beclin1 and the ratio of LC3II/LC3I were both significantly higher in the HG group than that in the control group. Beclin1 is an autophagy-related protein necessary for the formation of autophagosomes, which has increased expression in the retinas of diabetic rats.22 Arbutin treatment led to a further significant increase in the expression of beclin1 and in the ratio of LC3II/LC3I, which was lessened by addition of sirtinol (Figure 4C).
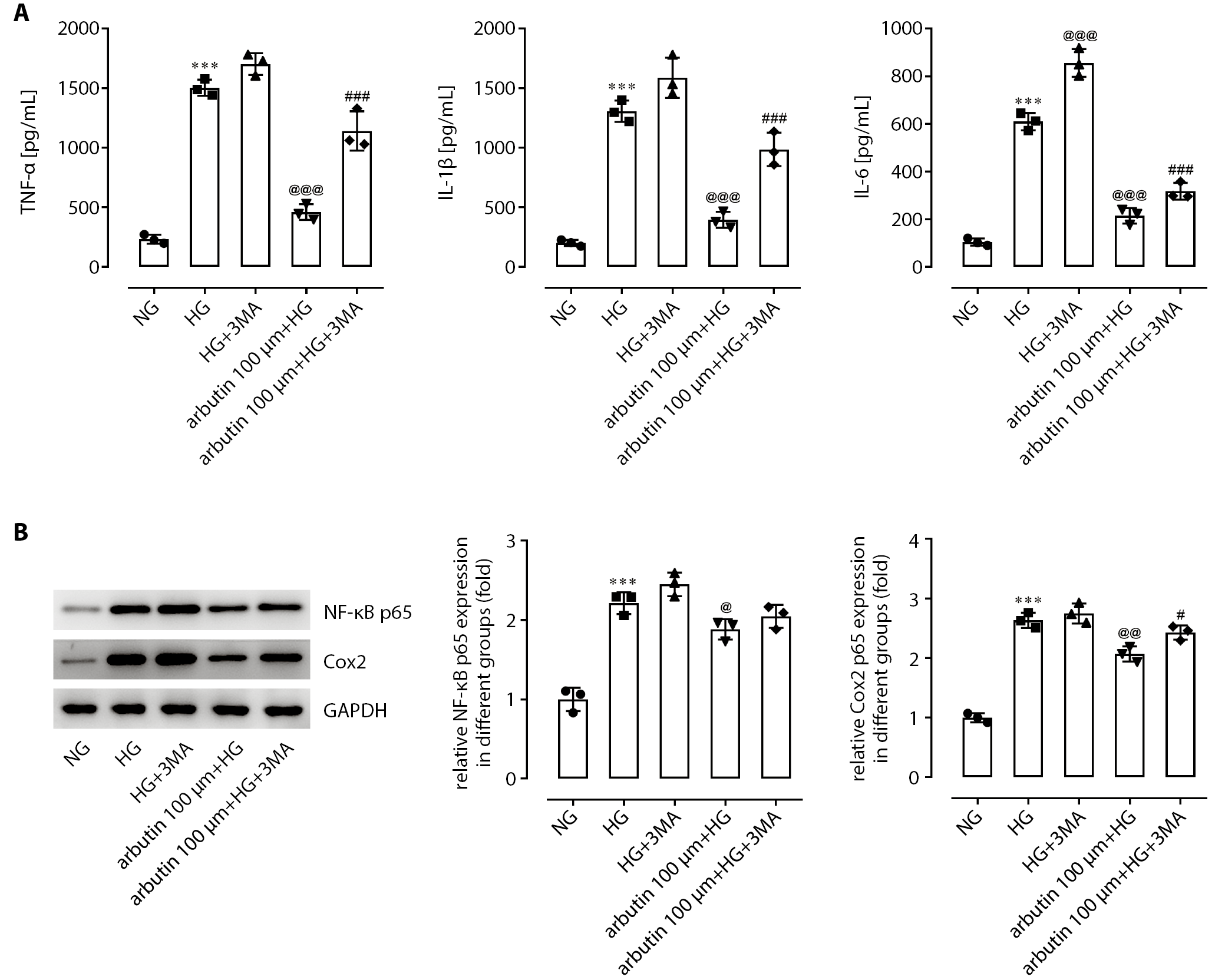
Autophagy blocking suppresses
the anti-inflammatory effects of arbutin
Following pretreatment for 1 h with 3-methyladenine (3MA), an autophagy inhibitor, and culture in a HG environment, the levels of TNF-a, IL-1β and IL-6 in cell supernatant were significantly higher than those cultured in a HG environment alone (Figure 5A). Moreover, 3MA pretreatment markedly blocked the effects of arbutin on these pro-inflammatory mediators, implying that the inflammation of APRE19 cells induced by HG may have been suppressed by arbutin via autophagy enhancement. Furthermore, we observed a similar action mechanism of arbutin in decreasing NF-KB p65 and COX-2 expression (Figure 5B).
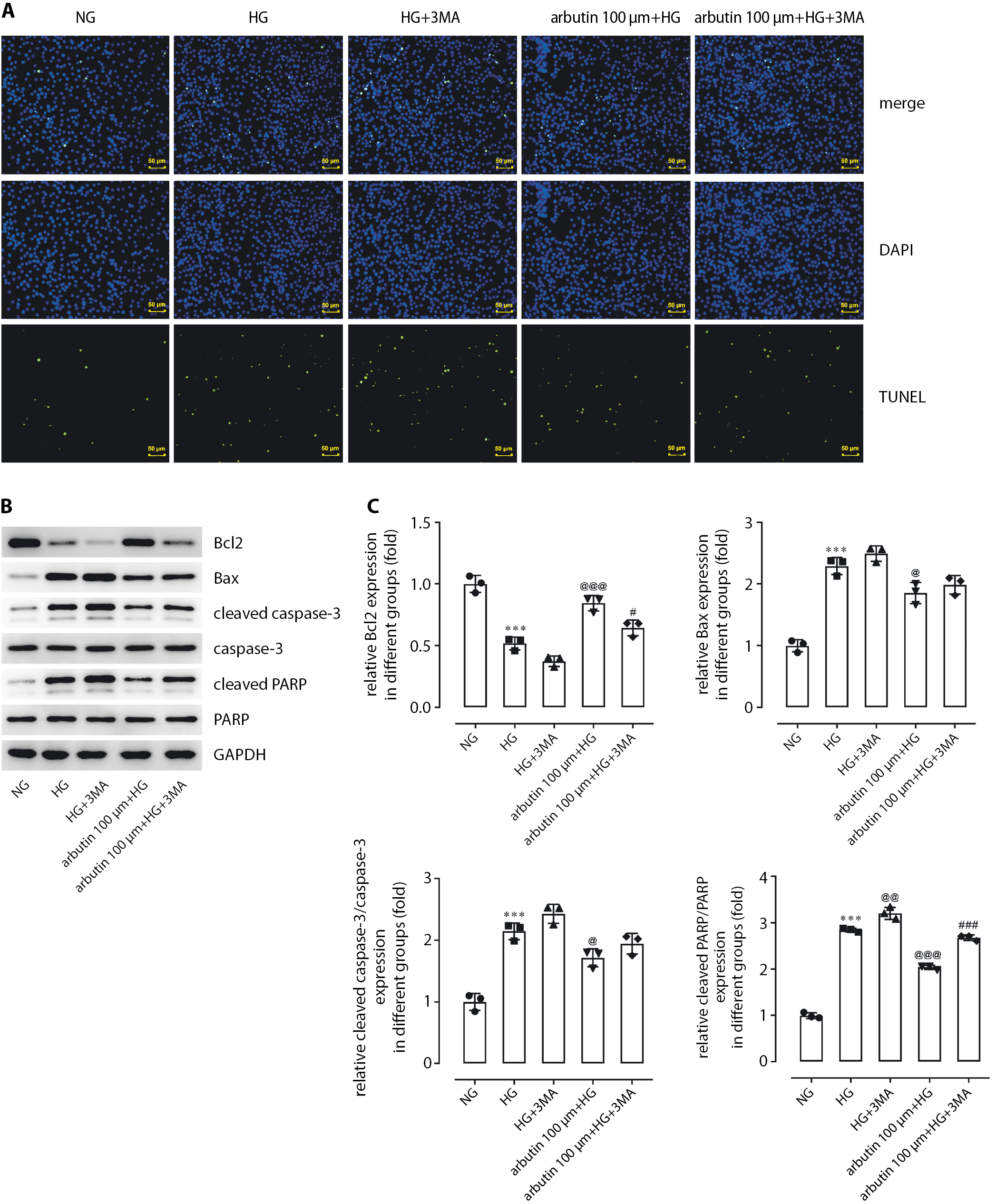
Inhibition of autophagy cancels out the anti-apoptotic effects of arbutin
Pretreatment with 3MA for 1 h increased the apoptosis level of APRE19 cells stimulated by HG in comparison with the HG group (Figure 6A). Expression levels of BAX, cleaved caspase-3 and cleaved PARP in the APRE19 cells were also elevated by 3MA while Bcl2 was decreased (Figure 6B,C). Furthermore, 3MA could reverse the influence of arbutin on the cellular behaviors brought about by HG induction, suggesting the involvement of autophagy in the inhibition of apoptosis by arbutin. Based on the abovementioned results, we drew the conclusion that arbutin produced inhibitory effects on inflammation and apoptosis by enhancing autophagy via SIRT1.
Discussion
Arbutin is commonly used as a protective agent, with no toxic effects to cells, and in vitro studies have shown that the activity of cells is not significantly affected by high concentrations of arbutin.23 Our findings are similar, in that ARPE19 cell viability was not significantly affected by an increase in arbutin concentration. Furthermore, arbutin markedly enhanced the viability of ARPE19 cells exposed to HG, suggesting that arbutin could protect these cells from HG-induced injury.
The presence of hyperglycemia in DR affects multiple biochemical pathways, such as the polyol, late glycosylated end-product and protein kinase C (PKC) pathways, the activation of which can lead to oxidative stress, inflammation and leucocyte stagnation, and induce the secretion of growth factors and cytokines, apoptosis and autophagy.20, 22, 24, 25, 26 Previous studies found that some inflammatory factors, such as IL-1β, IL-6 and TNF-α, are strongly correlated with the severity of DR.27, 28, 29
Inflammatory cytokines in RPE can be promoted in a HG environment,30 and high levels of TNF-α, IL-1β, and IL-6 were observed in APRE19 cells when induced by HG stimulation. These were significantly decreased by arbutin treatment in a dose-dependent manner. Hyperglycemia causes inflammation of the retina, which in turn leads to apoptosis.31 In addition to the inhibition of cell apoptosis by arbutin, the protein levels of Bcl2, BAX, cleaved caspase-3, and cleaved PARP were significantly downregulated in APRE19 cells treated with arbutin, indicating regulation of the apoptosis pathway, mediated by mitochondria. Cleaved PARP plays an important role in DNA damage repair and apoptosis, as a cleaving substrate of caspase and a core component of the apoptosis pathway.32, 33
The regulation of autophagy is closely connected to the progression of DR. Basal autophagy exerted anti-inflammatory effects, as recently reviewed.34 Under HG stimulation, increased autophagy levels may be insufficient to balance protein and organelle turnover.22, 34 We observed elevated autophagy levels in ARPE cells induced by HG, which were further enhanced by arbutin addition. It is surprising that sirtinol, as an inhibitor of SIRT1, significantly blocked the enhancement of autophagy by arbutin, although previously some studies have shown regulation of autophagy via SIRT1.
In our subsequent experiments, the inhibition of autophagy by 3MA could significantly hinder the regulatory effects of arbutin on HG-triggered ARPE19 cell behaviors that we confirm in this study. SIRT1 has been implicated in regulating inflammation and apoptosis processes in DR,35 and showed increased levels following arbutin treatment. The NFκB pathway regulates the expression of COX-2, through initiating transcription of the PTGS2 gene, which encodes COX-2.36 SIRT1 has been reported to suppress NF-κB activities, which are closely correlated with the expression of IL-1β, IL-6 and TNF-α, reduce COX-2 expression and play a vital role in inflammation.35, 37 Therefore, in the present study, arbutin could suppress inflammation by upregulating SIRT1 via inhibition of the NF-κB pathway. However, this needs to be confirmed by much deeper research. In RPE cells, SIRT1 could produce anti-apoptotic effects and reduce mitochondrial damage.38 Based on these experimental results, mitochondria-mediated apoptosis induced by HG could be suppressed through enhancement of SIRT1 expression by arbutin.
Limitations
There is still lack of in vivo evidence supporting the positive effects of arbutin on DR. Additionally, how arbutin promoted the expression of Sirt1 requires further deeper studies.
Conclusions
Arbutin treatment protected ARPE cells from inflammation and apoptosis induced by HG induction. The mechanism for this was autophagy enhancement via SIRT1. The present study provides novel insight into the treatment of DR and identifies a new molecular target for the development of therapy.