Abstract
Background. The 47-kDa membrane lipoprotein (Tp47) is the most representative membrane protein of Treponema pallidum (T. pallidum). Dendritic cells (DCs) are the most potent professional antigen-presenting cells (APCs) that connect innate and acquired immunity. The regulatory role of Tp47 on DCs remains unclear.
Objectives. To evaluate the effects of Tp47 on DC maturation and migration, and research the changes of the main chemokine C-C chemokine receptor type 7 (CCR7) involved in DC migration.
Materials and methods. A transwell assay was applied to assess the migration of DCs. Cytokines (interleukin (IL)-6, IL-10, IL-12, and tumor necrosis factor alpha (TNF-α)) in the supernatants were measured using enzyme-linked immunosorbent assay (ELISA), and the expression of cell surface markers (CD80, CD86, CD40, and human leukocyte antigen (HLA)-DR) and CCR7 was assessed using flow cytometry. The expression of CCR7 in DCs was analyzed using quantitative real-time polymerase chain reaction (qRT-PCR).
Results. The Tp47 promoted DC phenotypic maturation, such as increased CD40, CD80, CD86, and HLA-DR expression, as well as DC functional maturation, thus stimulating DCs to secrete inflammatory cytokines, including IL-6, IL-10, IL-12, and TNF-α. At the same time, Tp47 did not enhance DC migration and did not increase the expression of CCR7.
Conclusions. The Tp47 promoted the maturation of DCs while not enhancing CCR7-mediated DC migration ability. This may be one of the mechanisms by which T. pallidum escapes host immune clearance.
Key words: dendritic cells, migration, Treponema pallidum, syphilis, C-C chemokine receptor type 7
Background
Syphilis is a human infectious disease attributed to sexual transmission of the spirochete Treponema pallidum (T. pallidum). Its clinical manifestations are complex and can develop in stages. In the advanced stage, it can lead to multiple organ damage, seriously threatening the health of the host.1, 2 Previous studies have shown that T. pallidum can escape clearance by the human immune system and coexist with the host for a long time, which leads to asymptomatic and persistent infection, enabling sexual transmission of syphilis and causing physical and mental health problems.3, 4, 5 Different studies have suggested different mechanisms by which T. pallidum can evade immune control, thereby damaging host tissues. It is believed that antigenic variation and the paucity of outer membrane antigenic targets can promote the spread of T. pallidum.6, 7, 8, 9 In recent years, progressively more studies have suggested that host immune imbalance and immunosuppression are responsible for immune evasion of T. Pallidum.10, 11, 12
Dendritic cells (DCs), the most potent professional antigen-presenting cells (APCs), play an important role in the initiation and amplification of innate and adaptive immunity.13, 14, 15 The ability of DCs to migrate and activate naïve T cells is closely related to the efficiency of their immunomodulatory activity.16, 17, 18 Dendritic cells gradually mature after recognizing and ingesting antigens, and the expression of their surface molecule C-C chemokine receptor type 7 (CCR7) is upregulated. When CCR7 binds effectively to its ligand chemokine CCL2l/CCLl9, the resulting chemotaxis enables DCs to migrate into lymph nodes, triggering an adaptive immune response.19 The CCR7 is the major receptor on DCs to present antigens to naïve T lymphocytes, and it plays a key role in promoting the migration and nesting of DCs.20, 21, 22
Dendritic cells have been reported as one of the major immunocompetent cells that encounter bacterial antigens at primary sites of early syphilitic infection (e.g., mucous membranes or skin); they play a crucial role in clearing T. pallidum.23, 24 However, until now, there has been no study on the migratory capacity of DCs following T. pallidum infection. Previous studies have demonstrated that the outer membrane proteins and lipoproteins of T. pallidum constitute the main immune response components to T. pallidum.25, 26 Among them, the 47-kDa membrane lipoprotein (Tp47) is the most abundant membrane protein of T. pallidum. It has strong antigenicity and good specificity, mainly modulating the inflammation induced by T. pallidum.27, 28, 29, 30 Therefore, we chose Tp47 to induce a T. pallidum infection model in this study.
Objectives
This study aimed to evaluate the effects of Tp47 on the maturation and CCR7-mediated migration of DCs, which is expected to provide a preliminary explanation for why T. pallidum can escape clearance by the host immune system.
Materials and methods
Production of recombinant T. pallidum Tp47 protein after removal of endotoxin
Before the experiments, the recombinant T. pallidum Tp47 protein (0.6 g/L; Boson Biotech Co., Ltd., Xiamen, China) was treated to remove endotoxin contamination using an EtEraserTM endotoxin removal kit (Chinese Horseshoe Crab Reagent Manufactory Co., Ltd., Xiamen, China). The endotoxin content was detected using tachypleus amebocyte lysate (Chinese Horseshoe Crab Reagent Manufactory Co., Ltd.). The endotoxin content in the Tp47 preparation was limited to <0.05 endotoxin units/mL.
In vitro generation of dendritic cells
Through standard density gradient centrifugation using Ficoll-Paque™ PLUS (Cytiva, Marlborough, USA), human peripheral blood mononuclear cells (PBMCs) were isolated from 9 healthy blood donors (5 males and 4 females aged 18–55 years; all provided written informed consent). Then, cells from each donor sample were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, Waltham, USA) with 1% penicillin/streptomycin (Gibco) and 10% fetal bovine serum (FBS; Gibco), in a 6-well tissue culture plate, in a 37°C incubator with 5% CO2. Fresh medium containing 50 ng/mL recombinant human granulocyte-macrophage colony-stimulating factor (GM-CSF; R&D Systems, Minneapolis, USA) and 50 ng/mL recombinant interleukin (IL)-4 (R&D Systems) was replaced every 2 days. According to pre-experiments, we found that the confluence of DCs could reach 92.5 ±8.4% after human PBMCs were cultured in vitro in the presence of IL-4 and GM-CSF for 7 days.
On day 7, DCs were divided into 6 groups. The experimental group was stimulated with different concentrations of Tp47 (120, 300, 600, and 1200 ng/mL) based on previous studies.28, 30 Cells treated with 1 μg/mL lipopolysaccharide (LPS) or phosphate-buffered saline (PBS) were assigned to the positive control group or the negative control group, respectively. After 24 h of culturing, DCs in each group were collected for subsequent experiments.
Dendritic cell migration assay
Dendritic cell migration capacity was determined using a 5-μm pore size culture insert (Merck Millipore, Burlington, USA), which was placed into a 24-well culture plate as described elsewhere.20 Briefly, DCs in each group were harvested, washed twice with PBS and resuspended into a concentration of 1×106 cells/mL medium. Thereafter, 100 μL of DC suspension was seeded into the upper chamber, and 600 μL of medium complemented with 100 ng/mL CCL21 (PeproTech, Cranbury, USA) was added into the lower chamber. Three replicates for each cell type were placed in a 37°C, 5% CO2 incubator. At 1 h, 2 h, 3 h, and 4 h, 10 μL of the lower chamber culture, medium was obtained and counted on a hemocytometer under a light microscope (model CX33; Olympus Corp., Tokyo, Japan), and 10 μL of medium containing 100 ng/mL CCL21 was simultaneously added to the lower chamber. The DC migration rate was considered the ratio of migrated cells within the lower chamber to the total number of cells added to the upper chamber.
Cytokine assay
Using an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems) in accordance with the manufacturer’s instructions, the IL-6, IL-10, IL-12, and tumor necrosis factor alpha (TNF-α) concentrations in the DC culture supernatants were assessed. The absorbance was measured at 460 nm.
Flow cytometry
In each group, dendritic cells (1×105/mL) were resuspended in 100 µL PBS supplemented with 10 µg/mL fluorochrome-conjugated antibodies. The cells were stained with the following monoclonal antibodies: CCR7-PE (clone G043H7; BioLegend, San Diego, USA), CD86-APC (clone BU63; BioLegend), CD80-PE (clone 2D10.4; eBioscience, San Diego, USA), CD40-PE-Cy7 (clone 5C3; BioLegend), human leukocyte antigen (HLA)-DR-Alexa Fluor® 488 (clone LN3; BioLegend). After incubation with the antibodies in the dark for 30 min at 4°C, the cells were washed twice, resuspended in PBS (300 µL), and subjected to flow cytometry analysis (FACSCanto™ II; BD Biosciences, Franklin Lakes, USA).
Quantitative real-time polymerase
chain reaction
The expression of CCR7 on DCs was analyzed using quantitative real-time polymerase chain reaction (qRT-PCR). With the use of a TRIzol kit (Invitrogen, Waltham, USA), we extracted total RNA from the incubated cells. Using a Nanodrop™ 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, USA), RNA purity and concentration were tested. The RNA was reverse-transcribed into cDNA with the PrimeScript™ RT reagent kit (TaKaRa, Beijing, China), and amplification was conducted with TB Green Premix Ex Taq II (TaKaRa) in accordance with the manufacturer’s instructions. The qRT-PCR was performed with a PRISM 7300 Sequence Detection System (Applied Biosystems, Waltham, USA) using the following amplification conditions: 95°C for 30 s, followed by 40 cycles at 60°C for 1 min, 60°C for 34 s, 95°C for 15 s, 95°C for 5 s, and 95°C for 15 s. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) acted as an internal reference. Sangon Biotech Co., Ltd. (Shanghai, China) provided all primers. The primer sequences included: 5’-AGCAGCAGTGAGCAAGCGATG-3’ (forward) and 5’-GGGTGCGGATGATGACAAGGTAAC-3’ (reverse) for CCR7; and 5’-GCACCGTCAAGGCTAGAAC-3’ (forward) and 5’-TGGTGAAGACGCCAGTGGA-3’ (reverse) for GAPDH. The specificity of amplification was controlled using melting curve analysis. The 2−ΔΔCT method was adopted to analyze the data.31
Statistical analyses
Each experiment was repeated 3 times independently (n = 3). Variables were expressed as mean ± standard deviation (M ±SD), and the results were analyzed based on the “gemtc 1.4-2” package of R software (v. 4.1.2; R Foundation for Statistical Computing, Vienna, Austria). The statistical analysis was performed with the Wilcoxon–Mann–Whitney test, and the permutation test was employed to obtain the p-value. The Holm–Bonferroni method was used for post hoc corrections. A value of p < 0.05 was considered statistically significant. The full results (including p-values, test statistics and degrees of freedom, where appropriate) are available as Supplementary material for inspection (https://doi.org/10.5281/zenodo.7249173).
Results
Tp47 prompted DCs to secrete high levels of IL-6, IL-10, IL-12, and TNF-α
The maturation of DCs in response to pathogens mainly involves an increased expression of pro-inflammatory cytokines. As indicated by the ELISA results, the levels of IL-6, IL-10, IL-12, and TNF-α in the supernatants of cultured DCs in the presence of Tp47 (120, 300, 600, and 1200 ng/mL) were remarkably higher compared with those of the control group (Figure 1).
Tp47 upregulated the expression of CD80, CD86, CD40, and HLA-DR on DCs
In order to determine the effect of Tp47 on the phenotypic maturation of DCs, flow cytometry was carried out to detect the expression of surface molecules (CD80, CD86, CD40, and HLA-DR) on DCs. The results showed that the expression of surface molecules on DCs was upregulated under stimulation by Tp47 (120, 300, 600, and 1200 ng/mL) compared with the control group (Figure 2).
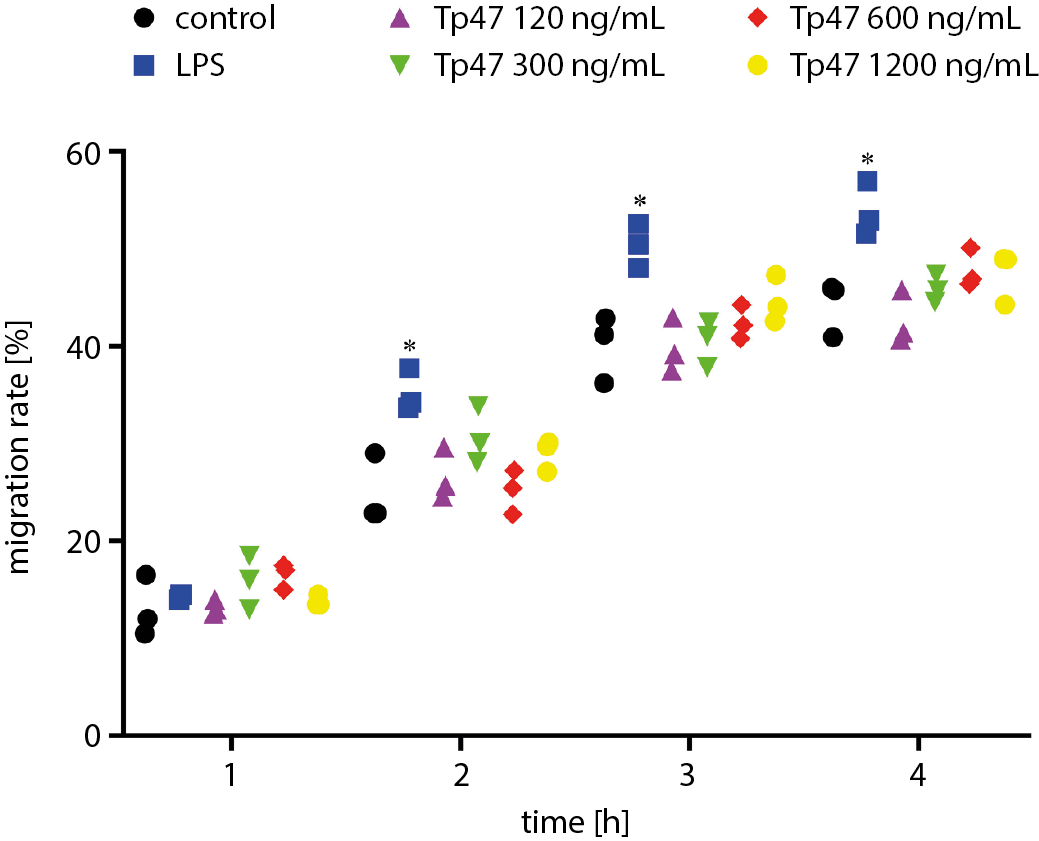
Tp47 did not promote DC migration
Using the transwell assay, we examined the migration ability of DCs in response to the chemokine CCL21. The results showed that the proportion of DCs that migrated to the lower chamber after 24 h of stimulation by Tp47 (120, 300, 600, and 1200 ng/mL) did not increase significantly compared with that of the control group. However, the proportion of DCs that, after stimulation with 1 μg/mL LPS, migrated to the lower chamber was significantly increased at 2 h, 3 h and 4 h compared to the control group (Figure 3).
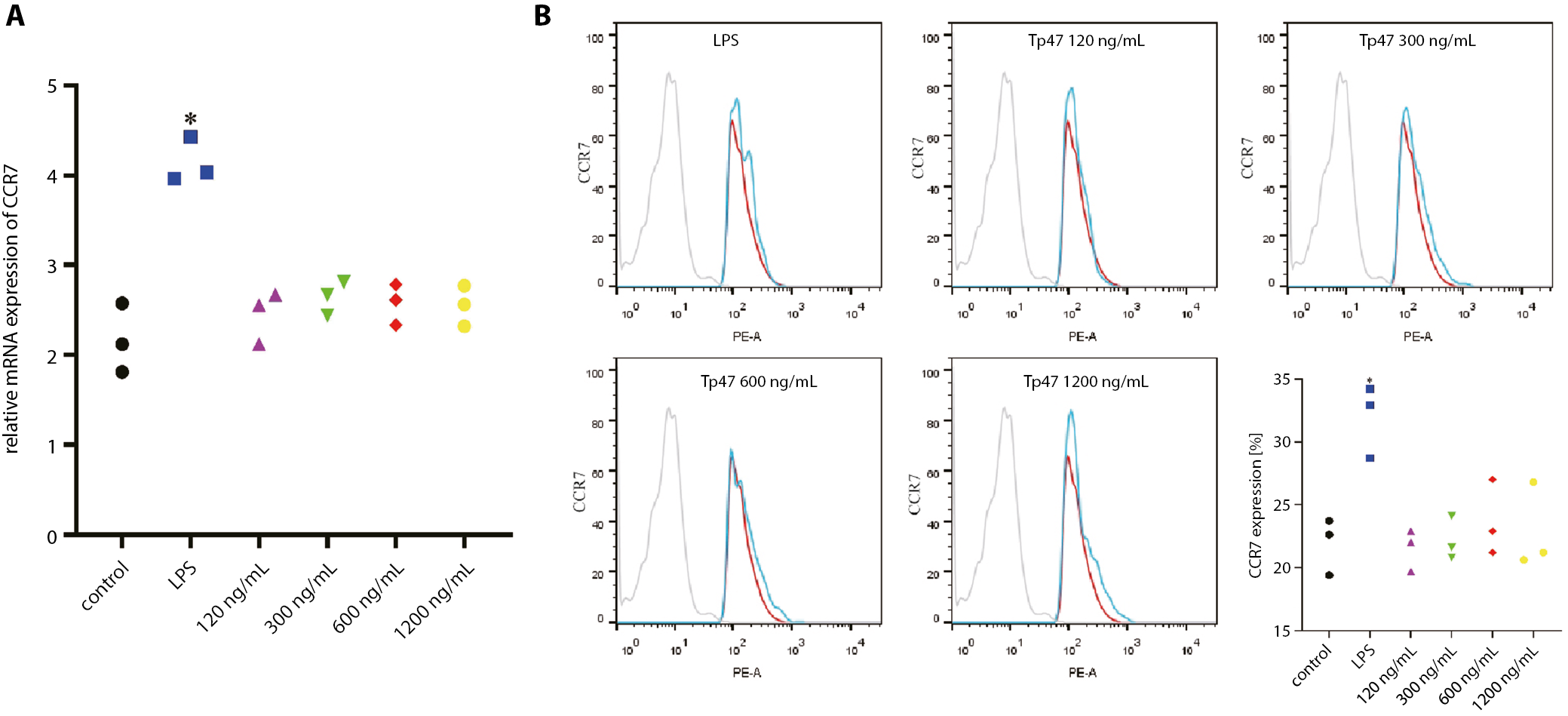
Tp47 did not increase CCR7 expression
Next, we verified whether CCR7 impacted DC migration after Tp47 stimulation. We assessed the CCR7 expression level in DCs using 2 approaches. First, the qPCR results showed that after co-culturing with Tp47 (120, 300, 600, and 1200 ng/mL), the expression level of CCR7 mRNA in DCs did not increase significantly compared with the control group (Figure 4A). Flow cytometry showed that the CCR7 level on DCs was not significantly upregulated after the stimulation with Tp47 at different concentrations in comparison with the control group (Figure 4B).
Discussion
Dendritic cells, as the most important APCs, have the ability to activate naïve T cells to generate adaptive immune responses only after maturation.32, 33 The maturation of DCs manifests as both phenotypic and functional.34 Phenotypic maturation implies that multiple surface molecules can be expressed at high levels, such as CD40, CD80, CD86, and HLA-DR, while functional maturation suggests the ability to secrete various inflammatory cytokines, including TNF-α, IL-6, IL-10, and IL-12.35 We found that for DCs stimulated by Tp47, not only the abovementioned surface molecule expression levels increased but also the ability of the DCs to secrete inflammatory cytokines significantly improved, indicating that Tp47 could simultaneously promote phenotypic and functional maturation of DCs. This result is consistent with previously reported studies on DCs in most pathogenic infections.36, 37, 38 After DCs uptake and process antigens, they undergo phenotypic and functional changes and gradually differentiate into mature DCs, which can provide the 2nd signal necessary for T-cell activation.39, 40
Immune cell migration serves as a key link between immune regulation and host defense. It has an indispensable role in eliminating the inflammatory responses triggered by foreign pathogens.41, 42 Previous studies have found that Tp47 can promote the migration and adherence of THP-1 cells to human dermal vascular smooth muscle cells by inducing the expression of monocyte chemoattractant protein-1 (MCP-1) and intercellular adhesion molecule-1 (ICAM-1).43 In our study, we explored the effect of Tp47 on migratory activity of DCs and found that the migratory ability of DCs did not significantly increase after they were stimulated by Tp47 compared to the control group. At the same time, we used Escherichia coli LPS stimulation as a positive control in all experiments. The results indicated that LPS could significantly promote the migration of DCs to the lower chamber in the transwell assay. Thus, it could be deduced that DCs did not fully exert their migratory ability to fulfill the mission of antigen presentation after Tp47 stimulation. During the maturation process, the migration of DCs to secondary lymphoid organs and their interactions with antigen-specific T cells are prerequisites for the induction of a primary immune response,44, 45 which plays an important role in anti-infective immunity. Numerous studies have shown that pathogenic microbes have evolved strategies to evade and/or suppress host inflammatory responses in order to infect and spread into other tissues and to other hosts. One of these strategies, as shown in Salmonella infection, is to inhibit DC migration.46, 47 In addition, Angeli et al. found that Schistosoma parasites inhibited DC migration to lymph nodes after infection through skin by producing prostaglandin D2.48 Our study indicated that Tp47, the most abundant T. pallidum protein, did not promote the migration of DCs, which might be one strategy by which T. pallidum evades host immune clearance.
It is well established that the interaction between CCR7 on the DC surface and its ligands has been proven to be a key driver for DC migration.49, 50 To further validate the impact of Tp47 on DC migration, we studied the change in the CCR7 expression level in DCs after Tp47 stimulation. The results showed that there was no increase in the DC surface CCR7 expression level and CCR7 mRNA content following Tp47 stimulation. Additionally, we found that LPS, as a positive control, significantly promoted the expression of CCR7. The chemotaxis jointly mediated by CCR7 and its ligands was the fundamental force driving DC migration that played an indispensable role in regulating the DC migration process. Studies have shown that the migration of DCs and the function of lymph nodes are significantly limited in the CCR7-knockout mice.51, 52 Our results reveal that Tp47 did not increase the CCR7 expression and, consequently, could not facilitate CCR7-mediated DC migration like LPS, which might be the mechanism underlying why Tp47 did not promote the migration of mature DCs. Interestingly, one of the important strategies utilized by pathogenic microorganisms to escape immune clearance is to modulate the CCR7 molecule on the surface of immune cells. Kling et al. found that the absence of CCR7 resulted in dysregulated monocyte migration and immunosuppression, facilitating chronic cutaneous leishmaniasis.53 Hartiala et al. reported that Borrelia garinii, one of the causative agents of Lyme borreliosis, impeded the migratory ability of DCs by significantly downregulating CCR7 expression in DCs.54 Therefore, Tp47, as previously confirmed by pathogenic microbes, could restrict the migration of DCs by reducing CCR7 expression to mitigate the impact of a robust immune response against itself.
Limitations
This study assessed the effects of Tp47 on DC maturation and migration. Despite the new findings presented here, our study has some limitations. First, immune evasion is a complex process that is not limited to the parameters evaluated above. Second, the suggested specific molecular mechanism underlying how Tp47 affects DC migration needs more studies to be confirmed. Finally, this study only included cell experiments in vitro with a small sample size, thus further confirmation of the findings with larger in vivo experiments is required.
Conclusions
In conclusion, the current findings indicate that the 47-kDa membrane lipoprotein of T. pallidum promoted the maturation of DCs without enhancing the CCR7-mediated DC migration. This study provides novel insights into the mechanisms by which T. pallidum escapes host immune clearance.
Supplementary files
The complete statistical results (including p-values, test statistics, and degrees of freedom where appropriate) are available as Supplementary material: https://doi.org/10.5281/zenodo.7249173.
Supplementary Table 1. Difference of IL-6 levels between other groups and the control group.
Supplementary Table 2. Difference of IL-10 levels between other groups and the control group.
Supplementary Table 3. Difference of IL-12 levels between other groups and the control group.
Supplementary Table 4. Difference of TNF-α levels between other groups and the control group.
Supplementary Table 5. Difference of CD40 levels between other groups and the control group.
Supplementary Table 6. Difference of CD80 levels between other groups and the control group.
Supplementary Table 7. Difference of CD86 levels between other groups and the control group.
Supplementary Table 8. Difference of HLA-DR levels between other groups and the control group.
Supplementary Table 9. Difference of CCR7 levels between other groups and the control group.
Supplementary Table 10. Difference of the relative CCR7 mRNA expression between other groups and the control group.
Supplementary Table 11. Differences of DCs migration rate between other groups and the control group at 1 h.
Supplementary Table 12. Differences of DCs migration rate between other groups and the control group at 2 h.
Supplementary Table 13. Differences of DCs migration rate between other groups and the control group at 3 h.
Supplementary Table 14. Differences of DCs migration rate between other groups and the control group at 4 h.