
















Abstract
Background. The regulatory effect of integrin β6 (ITGB6) on sweat gland cells in primary palmar hyperhidrosis (PPH) remains unclear.
Objectives. This study investigated the involvement of ITGB6 in the pathogenesis of PPH.
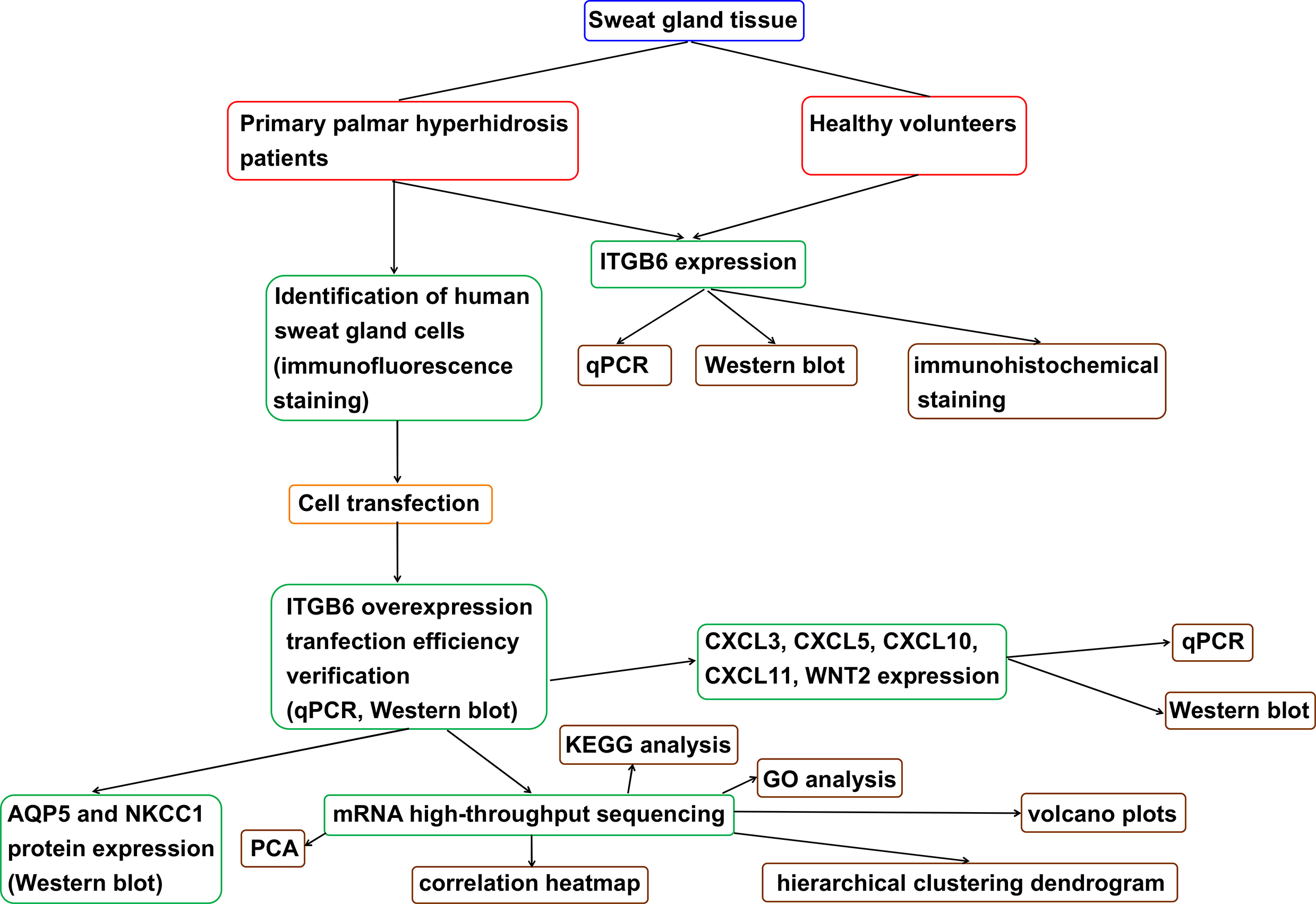
Materials and methods. Sweat gland tissues were collected from PPH patients and healthy volunteers. The expression levels of ITGB6 in sweat gland tissues were detected with quantitative polymerase chain reaction (qPCR), western blot and immunohistochemical staining. Sweat gland cells were extracted from PPH patients, and identified with immunofluorescence staining of CEA and CK7. The expression of aquaporin 5 (AQP5) and Na-K-Cl cotransporter 1 (NKCC1) in primary sweat gland cells that overexpress ITGB6 were also detected. Through a series of bioinformatic methods, differentially expressed genes in sweat gland tissues were examined and validated via comparing PPH samples and controls. The key proteins and biological functions enriched in PPH were determined using Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses.
Results. The ITGB6 was upregulated in sweat gland tissues of PPH patients compared to that of healthy volunteers. The CEA and CK7 were positively expressed in sweat gland cells extracted from PPH patients. The overexpression of ITGB6 upregulated AQP5 and NKCC1 protein expression in the sweat gland cells of PPH patients. A total of 562 differentially expressed mRNAs were identified using high-throughput sequencing (394 upregulated, 168 downregulated), which were mainly active in the chemokine and Wnt signaling pathways. After verification with qPCR and western blot, the overexpression of ITGB6 significantly upregulated CXCL3, CXCL5, CXCL10, and CXCL11, and downregulated Wnt2 mRNA and protein expression in sweat gland cells.
Conclusions. The ITGB6 is upregulated in PPH patients. It may be involved in the pathogenesis of PPH by upregulating AQP5, NKCC1, CXCL3, CXCL5, CXCL10, and CXCL11, and downregulating Wnt2 expression in sweat glands.
Key words: chemokines, ITGB6, AQP5, primary palmar hyperhidrosis, human sweat gland cells
Background
Sweat glands are important in adjusting body temperature in mammals.1 Hyperhidrosis is characterized by abnormally excessive sweating that far exceeds physiological needs. Palmar hyperhidrosis is the most common form of hyperhidrosis, which can be exacerbated by stress, heat, taste, and olfactory stimuli.2 It can seriously affect both the quality of life and work efficiency, and even cause severe psychological disorders if left untreated.3 At present, the main treatment of primary palmar hyperhidrosis (PPH) includes drug therapy, Traditional Chinese Medicine (TCM), direct current stimulation, and computed tomography (CT)-guided percutaneous puncture thoracic sympathetic blockade.4 However, most therapeutic strategies are limited to the relief of symptoms rather than a cure. Therefore, clarifying the molecular mechanism of PPH and developing target drugs are crucial.
As a cell surface receptor, integrins are transmembrane heterodimers composed of an α subunit and a β subunit that mediate signal transduction between the cells and the extracellular matrix.5 Integrin β6 (ITGB6) belongs to the integrin family and is only expressed in epithelial tissues, having important roles in regulating wound healing, inflammatory responses and tumorigenesis. Wu et al. reported impaired wound repair in ITGB6−/− mice with corneal debridement and keratectomy wounds.6 Moreover, Chen et al. demonstrated that the overexpression of ITGB6 in intestinal epithelial cells promotes dextran sulfate sodium (DSS)-induced colitis in mice.7 Zheng et al. found that ITGB6 silencing can inhibit the progression of cervical cancer by modulating the JAK/STAT3 signaling pathway.8 To date, the regulatory effect of ITGB6 on sweat gland cells in PPH remains uncertain.
Objectives
The aims of this study were to analyze the expression of ITGB6 in sweat gland tissue obtained from patients with PPH at the clinical level, explore the possible mechanism of ITGB6 in regulating the function of human sweat gland cells through the isolation of primary human sweat gland cells and the construction of ITGB6 overexpression vector, and analyze the underlying mechanism of ITGB6 participation in the pathogenesis of PPH. The findings of this study may serve as guidance for the treatment of PPH.
Materials and methods
Ethics approval
The Ethics Committee of The First Affiliated Hospital of Fujian Medical University (Fuzhou, China) approved this study (approval No. [2019]261). Written informed consent was obtained from all examined patients before sample collection.
Sample collection
Axillary all-layer skin samples (2 × 5 mm) of 8 patients with PPH treated with single-port surgery at the natural folds of the axilla in The First Affiliated Hospital of Fujian Medical University from January to September 2019 were collected as the experimental group. The samples were obtained from the right axilla by incision. Patients with a history of secondary hyperhidrosis, such as hyperthyroidism and tuberculosis, were excluded. All-layer skin samples (2 × 5 mm) from the right axilla of patients without hyperhidrosis (n = 11) who underwent axillary plastic surgery were surgically collected as the control group. The samples were obtained by incision. The patients had no history or family history of palmar and axillary hyperhidrosis and axillary osmidrosis.
Reagents and instruments
We used the following primary antibodies in our study: rabbit anti-ITGB6 antibody (19695-1-AP) was obtained from Proteintech (Wuhan, China), while rabbit anti-aquaporin 5 (AQP5) antibody (AF5169), rabbit anti-Na-K-Cl cotransporter 1 (NKCC1) antibody (DF2245), rabbit anti-CEA antibody (BF0092), rabbit anti-CK7 antibody (AF0195), rabbit anti-CXCL3 antibody (DF8554), rabbit anti-CXCL5 antibody (DF9919), rabbit anti-CXCL10 antibody (DF6417), rabbit anti-CXCL11 antibody (DF9917), and rabbit anti-WNT2 antibody (DF8067) were all purchased from Affinity Biosciences (Cincinnati, USA). The mouse monoclonal anti-GAPDH antibody (TA-08) and horseradish-labeled goat anti-rabbit IgG (H+L) antibody (ZB-2301) were both obtained from ZSGB-Bio (Beijing, China). The DAB Chromogenic Substrate Kit (CW0125), UltraSYBR Mixture (CW0957M), neutral resin (CW0136), BCA Protein Assay Kit (CW0014S), TRIzol reagent (CW0580S), UltraPure mRNA Purification Kit (CW0581M), and HiFiScript cDNA Synthesis Kit (CW2569M) were all purchased from CWBIO (Beijing, China). The hematoxylin and eosin (H&E) staining kit (AR1180-1) was purchased from Boster Bio (Pleasanton, USA), Opti-MEM (31985-062) from Gibco (Grand Island, USA), and radioimmunoprecipitation assay (RIPA) lysis buffer (C1053) from Applygen (Beijing, China).
Cell culture incubator (BPN-80CW) was purchased from Shanghai Yiheng Technology (Shanghai, China), and the bench-top low-temperature high-speed centrifuge (5424R) from Eppendorf (Hamburg, Germany). The CFX Connect™ real-time fluorescence PCR instrument was manufactured by Bio-Rad (Shanghai, China), and the vertical electrophoresis apparatus (DYY-6C) and microplate reader (WD-2102B) by Beijing Liuyi Biotechnology Co., Ltd. (Beijing, China). The automatic high-speed grinding device (Tiss-12) was obtained from Shanghai Jingxin Biological Pharmaceutical Co. Ltd. (Shanghai, China) and the ChemiDocTM XRS+ System from Bio-Rad. Fluorescence microscopes (model CX41 and model CKX53) were purchased from Olympus Corp. (Tokyo, Japan), the multifunctional enzyme labeling analyzer (Safire II) from Tecan (Männedorf, Switzerland), and the manual rotary microtome (2235) from Leica Biosystems (Wetzlar, Germany). The Nanodrop 2000 was obtained from Thermo Fisher Scientific (Waltham, USA), Agilent 2100 from Agilent Technologies (Santa Clara, USA), and the HiSeq4000 sequencing platform from Illumina (San Diego, USA).
Extraction of sweat gland cells
The human sweat gland tissues were collected, immersed in 75% ethanol for 10 s, and washed in phosphate-buffered saline (PBS) containing 1% penicillin and streptomycin. The apocrine sweat glands were isolated under a microscope and then incubated in PBS. The outer membrane of the apocrine sweat glands was peeled off, the remaining tissues were cut off and centrifuged for 4 min at 1000 rpm. The mixture was digested in 2 g/L collagenase II overnight, and for 10 min at 37°C the following day. After the termination of the enzymatic digestion, the mixture was centrifuged for 4 min at 1000 rpm, the supernatant was discarded, and freshly prepared Dulbecco’s modified Eagle’s medium/Nutrient Mixture F-12 (DMEM/F12) + 20% fetal bovine serum (FBS) + 1% penicillin and streptomycin were added, inoculated in a culture flask and placed in 37°C with 5% CO2 for culture.
The cell culture supernatant was discarded, and cells were washed twice in 1× PBS. Then, 0.25% trypsin containing 0.02% ethylenediaminetetraacetic acid (EDTA) was added. Cell digestion was terminated by adding the culture medium when the cell morphology became round. Then, the cell suspension was collected into a 10-milliliter centrifuge tube and centrifuged for 3 min at 1000 rpm. The supernatant was discarded, and the culture medium was added and resuspended evenly. The cell suspension was harvested into the culture plate and cultured in a 37°C, 5% CO2 incubator.
Immunohistochemical staining
Paraffin-embedded tissue sections of sweat glands were dewaxed, incubated in citrate buffer for antigen retrieval, heated for 2 min in a pressure cooker, and then naturally cooled to room temperature. After washing with PBS, the sections were put in a wet box at room temperature, where newly prepared 3% H2O2 was added to quench endogenous peroxidase activity. Ten minutes later, the sections were washed in PBS for 5 min (3 times), and any excess reagent was wiped away. After blocking with 5% bovine serum albumin (BSA) at 37°C for 30 min, samples were incubated with the anti-ITGB6 antibody at 4°C overnight, followed by secondary antibodies at 37°C for 30 min. Then, the sections were stained with 3,3’-diaminobenzidine (DAB) solution, counterstained with hematoxylin, washed in tap water, mounted, and observed under a microscope (model CX41; Olympus Corp.).
Immunofluorescence
Cells cultured in dishes were washed in PBS 3 times for 3 min each, fixed in 4% paraformaldehyde for 15 min and rewashed in PBS 3 times for 3 min each. After immersion in PBS containing 0.5% Triton X-100 for 20 min at room temperature, the cells were washed in PBS 3 times for 5 min, and the medium was cleared. Then, the cells were blocked in 5% BSA for 30 min at 37°C and incubated with anti-CEA and anti-CK7 antibodies for 3 h at 37°C. Following 3 washes with PBS for 3 min each, the samples were incubated with the appropriate secondary antibody at 37°C in the dark for 45 min and washed 3 times in PBS for 3 min each. Then, the cells were incubated with 4’,6-diamidino-2-phenylindole (DAPI) for 5 min for nuclear counterstaining and washed in PBS. Slides were sealed with 50% glycerol, followed by examination under a fluorescence microscope (model CKX53; Olympus Corp.) in the dark.
Cell transfection
The cells cultured to 70% density were prepared for transfection by replacing the media with serum-free medium. Two sterilized Eppendorf tubes were prepared with equal volumes of Opti-MEM. Then, 5 µL of Lipofectamine 3000 were added to 1 tube, while 2.5 µg of plasmid and 5 µL of P3000 were added to another tube, mixed well and incubated for 5 min at room temperature. Then, the 2 tubes were evenly mixed and incubated for 15 min at room temperature. The mixed solution was applied to a 6-well plate and cultured in the incubator. Four hours after the transfection, 1 mL of complete medium containing 20% serum was added, and verification was performed after 48 h.
High-throughput sequencing
After preparation of the cDNA library, total RNA was extracted using TRIzol reagent, and the concentration and purity were detected using a Nanodrop 2000. The RNA integrity was determined with gel electrophoresis. The RNA integrity number (RIN) value was measured using Agilent 2100. The total amount of RNA required for single library construction was >5 µg, the concentration was ≥200 ng/μL, and optical density (OD) 260/280 was between 1.8 and 2.2. The Ribo-Zero Magnetic kit (EpiCentre, Madison, USA) was used to remove rRNA, and RNase R (EpiCentre) was used to remove linear RNA. The TruSeqTM Stranded Total RNA Library Prep Kit (Illumina) was used to construct the paired-end sequencing library. The HiSeq4000 sequencing platform was used for sequencing and data analysis, and SepPrep (https://github.com/jstjohn/SeqPrep) and Sickle (https://github.com/najoshi/sickle) software were used to determine the quality of data. Qualified data were compared and analyzed with the reference genome data using the bowtie method.
Western blot
The samples from each group were added to the corresponding lysis buffer, lysed on ice for 20 min and centrifuged at 12,000 rpm for 10 min. The supernatant was carefully aspirated to obtain the total protein, and the protein concentration analysis was performed using a bicinchoninic acid (BCA) kit. The protein was denatured, loaded and subjected to sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) for 1–2 h, followed by wet transfer to a polyvinylidene fluoride (PVDF) membrane for 30–50 min. The incubation with the primary antibody solution was conducted at 4°C overnight, followed by the secondary antibody at room temperature for 1–2 h. Then, the enhanced chemiluminescence (ECL) exposure solution was dropped on the membrane and imaged in a gel imaging system. The grey value of any bands that were present was analyzed with Quantity One software (Bio-Rad).
Quantitative polymerase chain reaction
The samples were collected, and TRIzol lysis buffer was added and mixed thoroughly with a pipette gun to ensure that the adherent cell suspension was in full contact with the lysis buffer. Then, the cell suspension was collected to extract the total RNA. The RNA was synthesized using reverse transcription HiFiScript first-strand cDNA synthesis kit. Fluorescence quantitative polymerase chain reaction (qPCR) was performed. The reaction system, steps and primer sequences for qPCR are presented in Table 1. Primers were synthesized by General Biol (Anhui) Corp. Ltd. (Chuzhou, China). The β-actin was used as an internal reference, and the relative expression was calculated according to the 2−ΔΔCT method.
Statistical analyses
The IBM SPSS v. 20.0 software (IBM Corp., Armonk, USA) was used for statistical analysis. All experiments were performed 3 times, and the quantitative results were expressed as mean ± 95% confidence interval (95% CI). Quantitative comparison between the 2 groups was performed using independent samples t-test (Student’s t-test). When comparing multiple groups, a quantitative numerical comparison was performed with one-way analysis of variance (ANOVA) and post hoc Tukey’s honest significant difference (HSD) test. The p-value of the test level was α = 0.05, and p < 0.05 indicated a significant difference.
Principal component analysis
Principal component analysis (PCA) is often used to reduce the dimensionality of a dataset while maintaining the features that contribute the most to the variance, thereby effectively finding the main elements and structures in the data, and removing noise and redundancy. In addition, the original complex data are dimensionally reduced to reveal the simple structure hidden behind the complex data. By analyzing the components of different sample species and functions, the differences and distances between samples can be discovered. Principal component analysis uses variance decomposition to reflect the differences between multiple sets of data on the coordinate axis. For example, the more similar the sample composition, the closer the distance is reflected in the PCA diagram. Samples between different environments may show dispersive and aggregated distributions. The closer the samples are to different environments, the more likely they are to show a dispersive and agglomerated distribution, and the 2 or 3 components with the highest explanatory power for sample differences in PCA results can be used to verify hypothetical factors.
Volcano plot
A volcano plot is a type of scatterplot that shows statistical significance (p-value) compared to magnitude of change (fold change), and enables quick visual identification of genes with large fold changes that are also statistically significant. These may be the most biologically significant genes.
Gene Ontology analysis and Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis
Gene Ontology (GO) is an international standardized gene function classification system that provides a set of dynamically updated standard vocabulary (controlled vocabulary) to comprehensively describe the attributes of genes and gene products in organisms. Gene Ontology has 3 ontologies in total, which describe the molecular function, the cellular component and the biological process involved in the gene. The basic unit of GO is a term, and each term has a unique identifier (composed of “GO” plus 7 numbers, such as GO: 0072669). The terms of each type of ontology form a directed acyclic topology through their connections (is_a, part_of, regulate). The horizontal axis is the functional classification, and the vertical axis is the number of genes in the classification (right) and its percentage in the total number of annotated genes (left). Different colors represent different classifications. Light color represents target genes, and dark color represents all genes.
Kyoto Encyclopedia of Genes and Genomes (KEGG) is a comprehensive database of biological systems, integrating genomic, chemical and system functional information. Among them, KEGG GENES collects the gene protein sequences of all known complete genomes, including the minimum information on each gene. The KEGG ORTHOLOGY (KO) system links various KEGG annotation systems together. Kyoto Encyclopedia of Genes and Genomes has established a complete KO annotation system, which can complete the functional annotation of the genomes or transcriptome of newly sequenced species.
Results
Demographic and clinical data
The control group included 3 males and 8 females, with a mean age of 21.8 ±5.0 years. The experimental group consisted of 2 males and 6 females, with a mean age of 21.4 ±5.3 years. All patients were of Han ethnicity with no previous history of surgical treatments. Both groups were comparable in terms of gender, mean age, ethnic background, and history of surgical treatments.
ITGB6 is upregulated in sweat gland tissue of patients with primary palmar hyperhidrosis
Compared with the healthy volunteers, both mRNA and protein expressions of ITGB6 in the sweat gland tissues of patients with PPH were significantly higher (Figure 1A,B). Immunohistochemical staining consistently demonstrated a significantly increased staining of ITGB6 in the sweat gland tissue of patients with PPH compared to the controls (Figure 1C).
Identification of human sweat gland cells and verification of ITGB6 overexpression transfection efficiency
The CEA and CK7 are generally considered markers of sweat gland cells,9 and both CEA and CK7 proteins were expressed in primary isolated human sweat gland cells (Figure 2A). The mRNA and protein expression of ITGB6 in cells transfected with ITGB6 overexpression (OE) were significantly upregulated compared to those transfected with OE negative control (NC), suggesting high transfection efficacy (Figure 2B–D).
Regulatory effects of ITGB6 on aquaporin 5 and Na-K-Cl cotransporter 1 protein expression levels in sweat gland cells
Aquaporin 5 is a water channel protein that participates in the generation of saliva, tears and pulmonary secretions. Moreover, NKCC1 is vital for organs that secrete fluids. Therefore, we explored the influence of ITGB6 on AQP5 and NKCC1 protein expression levels in sweat gland cells extracted from patients with PPH using western blot. Interestingly, the overexpression of ITGB6 upregulated the AQP5 and NKCC1 protein expression levels (Figure 3A,B).
mRNA high-throughput sequencing analysis
We found large differences in gene expression levels between the ITGB6 OE group and the control group (Figure 4A–C). In Figure 4D, the horizontal axis is the fold change (log (B/A)) value of the difference of transcript expression between samples of different groups, and the vertical axis is the p-value that represents the statistical significance of the expression changes. Compared with the OE NC group, a total of 562 differentially expressed mRNAs in the ITGB6 OE group were determined with high-throughput sequencing, involving 394 upregulated and 168 downregulated mRNAs. They were mainly enriched in the chemokine and Wnt signaling pathways. The results of GO and KEGG pathway enrichment analyses are presented in Figure 5A and Figure 5B, respectively.
Regulatory effects of ITGB6 on expression levels of CXCL3, CXCL5, CXCL10, CXCL11, and WNT2 in sweat gland cells
We further examined the expression of CXCL3, CXCL5, CXCL10, CXCL11, and WNT2 in sweat gland cells extracted from patients with PPH and transfected with OE NC or ITGB6 OE. We found that CXCL3, CXCL5, CXCL10, and CXCL11 mRNA and protein expression levels were significantly upregulated, whereas WNT2 was significantly downregulated in sweat gland cells overexpressing ITGB6 (Figure 6A–C).
Discussion
Primary palmar hyperhidrosis refers to abnormal hypersecretion of sweat glands in the hands, which is not related to the temperature of the external environment. It manifests as wet palms in mild cases, while those severely affected may drip sweat from their hands, posing great psychological pressure and disrupting their social life.10, 11 At present, treatment measures can only alleviate symptoms rather than cure the disease,12 and thus, it is of great importance to explore the pathogenesis of PPH and develop targeted interventions. In the present study, ITGB6 was found to be upregulated in sweat gland tissues of patients with PPH compared to healthy volunteers. Furthermore, it was shown that ITGB6 OE may upregulate AQP5 and NKCC1 in sweat gland cells extracted from patients with PPH. Combined with mRNA high-throughput sequencing and verification experiments, it was found that ITGB6 OE can significantly increase the CXCL3, CXCL5, CXCL10, and CXCL11 mRNA and protein levels while inhibiting WNT2 mRNA and protein expression.
As the main component of sweat, water molecules are regulated by relevant proteins expressed on the cell membrane of sweat glands. The AQP5 is a water channel protein that specifically transports water across the membrane. It is involved in the secretion and absorption of water and balances intra- and extracellular water through increasing water permeability of the cell membrane.13 In recent years, studies have reported that AQP5 participates in various physiological functions of human water regulation. Wang et al. found that the inhibition of HMGB1 could alleviate the symptoms of Sjögren’s syndrome by inhibiting the HMGB1/TLR4/NF-κB signaling pathway and upregulating the expression of AQP5.14 Furthermore, Du et al. reported that AQP5 is highly expressed in the axillary sweat glands of patients with primary focal hyperhidrosis.15 Our previous study demonstrated that ACVR1 can promote human sweat gland cell proliferation by upregulating AQP5 expression.16 Finally, the downregulation of CHRNA1 can alleviate the symptoms of PPH by inhibiting AQP5 expression.17 In the present study, ITGB6 OE may upregulate the expression of the AQP5 protein in sweat gland cells extracted from patients with PPH, suggesting that ITGB6 may aggravate the progression of PPH by regulating the expression of aquaporin. We found a similar result for NKCC1, whose main function is to transport Na, K and Cl ions into sweat gland cells in the ratio of 1:1:2. It plays an important role in cell ion transport, stabilizing ion gradient and regulating cell volume.18, 19 Our previous study found that ACVR1 can promote sweat gland cell proliferation by upregulating NKCC1 expression, and its main mechanism of action may be achieved by regulating the transport of Na, K and Cl ions.16
Chemokines contribute to the fight against and clearance of foreign bodies like invading pathogens through the directional chemotaxis of immune cells.20, 21, 22 According to the differences in cysteine in the chemokines, they can be divided into 4 main subfamilies: CXC, CC, CX3C, and XC. Among them, the CXC subfamily, as a research hotspot, plays an important role in various diseases.23 Pro-inflammatory factors, CXCL3, CXCL5, CXCL10, and CXCL11 have been reported to play key regulatory roles in various diseases such as acute lung injury,24 atherosclerosis25 and ovarian cancer.26 However, their potential involvement in PPH is unclear. Our bioinformatic analysis revealed that CXCL3, CXCL5, CXCL10, and CXCL11 were upregulated in PPH with ITGB6 OE. As a result, we speculate that ITGB6 exacerbates the progression of PPH by promoting the accumulation of neuroinflammatory factors during sweat gland secretion.
The Wnt signaling pathway is highly conserved across species and exists widely in multicellular organisms.27 It is capable of regulating cell proliferation, differentiation, apoptosis, migration, and invasion, and plays an important role in the occurrence and development of various diseases.28 Cui et al. reported that the Wnt signaling pathway participates in the regulation of the induction and development of sweat glands.29 Therefore, we speculated that ITGB6 promoted the progression of PPH by inhibiting the activation of Wnt signaling in the process of sweat gland secretion. Our results showed that ITGB6 OE downregulated WNT2 in sweat gland cells extracted from patients with PPH.
Limitations
This study mainly explored the potential mechanism of ITGB6 in the pathogenesis of PPH. Further studies may be needed to investigate the involved mechanism more deeply.
Conclusions
The ITGB6 was upregulated in patients with PPH, and the overexpression of ITGB6 may promote AQP5 and NKCC1 protein expression. Moreover, ITGB6 OE significantly increased CXCL3, CXCL5, CXCL10, and CXCL11 mRNA and protein expression, while inhibiting WNT2 mRNA and protein expression.
Supplementary data
The Supplementary materials are available at https://doi.org/10.5281/zenodo.7750959. The package contains the following files:
Supplementary Table 1. Data analysis for Figure 1A–C, Figure 2B,D, Figure 3B, Figure 4A–C, and Figure 5A,C.
Supplementary Table 2. Data analysis for PCA rotation (Figure 4A).





