
















Abstract
Background. Hepatic progenitor cells (HPCs) play an important role in the treatment of chronic liver disease.
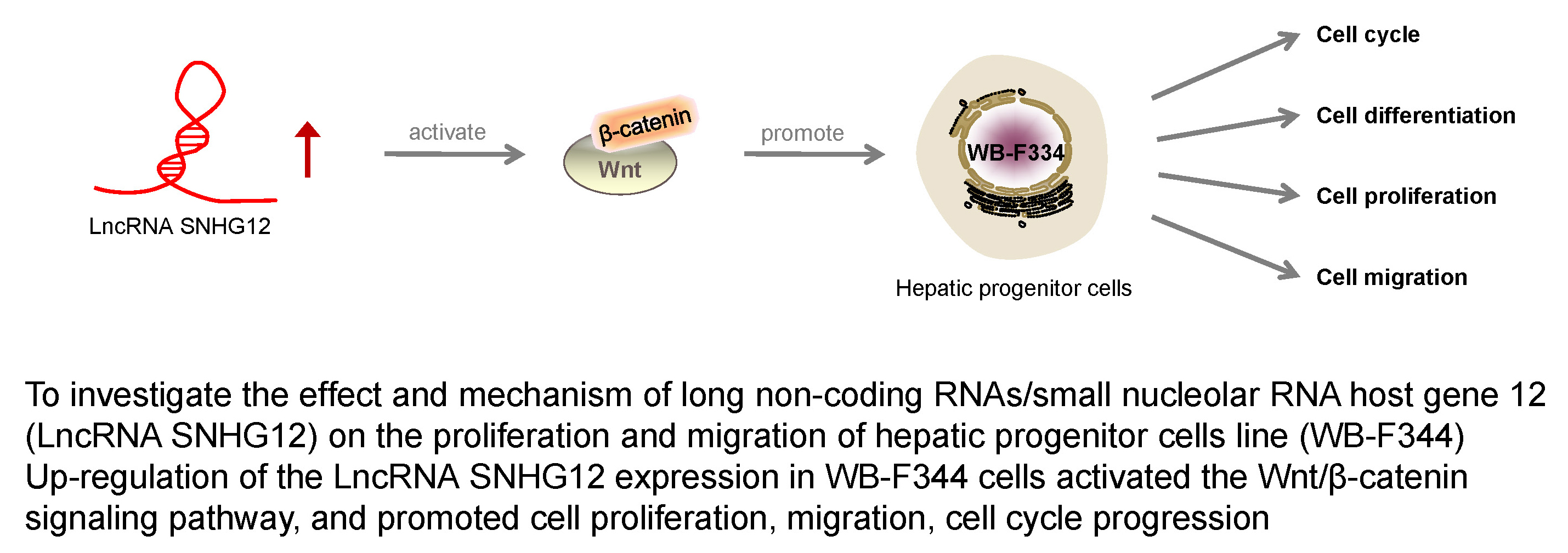
Objectives. To investigate the effect and mechanism of long noncoding RNAs/small nucleolar RNA host gene 12 (lncRNA SNHG12) on the proliferation and migration of the HPC cell line WB-F344.
Materials and methods. Hepatic progenitor cells were divided into a no-treatment group (sham), empty vector transfection of plasmid pcDNA3.1 (NC vector), pcDNA3.1-SNHG12 (SNHG12), negative short hairpin RNA (sh-NC), SNHG12 shRNA (sh-SNHG12), and pcDNA3.1-SNHG12+salinomycin intervention (SNHG12+salinomycin) groups. Cell proliferation, cell cycle and migration ability, as well as albumin (ALB), alpha-fetoprotein (AFP), β-catenin, cyclin D1, and c-Myc protein expression in each group were determined using Cell Counting Kit-8 (CCK-8), 5-ethynyl-2’-deoxyuridine (EdU), flow cytometry, transwell migration assays, enzyme-linked immunosorbent assay (ELISA), and western blot.
Results. The overexpression of lncRNA SNHG12 significantly upregulated proliferation, migration and cell cycle progression of WB-F344 cells. Furthermore, the overexpression of lncRNA SNHG12 increased the level of ALB, and the protein expression of β-catenin, cyclin D1 and c-Myc in the cell line, while decreasing the level of AFP. Conversely, the knockdown of lncRNA SNHG12 displayed the opposite effects. The inhibition of the Wnt/β-catenin signaling pathway with salinomycin significantly downregulated the β-catenin, cyclin D1 and c-Myc protein expression in WB-F344 cells.
Conclusions. The lncRNA SNHG12 promotes the proliferation and migration of WB-F344 cells via activating the Wnt/β-catenin pathway.
Key words: proliferation, migration, Wnt/β-catenin, WB-F344, lncRNA SNHG12
Background
With its complex structure and variety of biological functions, the liver is one of the most important organs in vertebrates.1 Specifically, its biological functions include synthesis and decomposition of liver glycogen, synthesis and transportation of lipids, biosynthesis of hormones and proteins, as well as drug metabolism and the decomposition of harmful components.2 Furthermore, there is a close link between the liver and other tissues, including skeletal muscle, adipose tissue, brain, and other viscera.3 Hence, liver injury or functional impairment may be life-threatening. To this end, organ transplantation seems to be the only option for the most critical liver diseases. However, the lack of liver donors restricts the treatment of many patients who need liver transplants.4, 5 Therefore, hepatic progenitor cells (HPCs) have become a relevant research avenue because of their potential to replace liver transplant operations. Hepatic progenitor cells, also known as small hepatocyte or biliary epithelioid cells, are a heterogeneous cell type with bidirectional differentiation potential.6 Studies have shown that HPCs play a vital role in the development of liver regeneration and liver cancer, which can be applied in the treatment of chronic liver diseases.7, 8 Najimi et al. transplanted human-derived HPCs into severe combined immunodeficient mice through the splenic vein. They found that HPCs could proliferate and differentiate into hepatocytes after transplantation, indicating that they could be used as a replacement or adjuvant therapy for the treatment of liver diseases.9 Moreover, HPCs can be obtained from the patients themselves and the population expanded in vitro, thereby preventing the immune rejection caused by allogeneic transplantation. Therefore, HPC transplantation is expected to be another treatment option alongside liver tissue transplantation.9 However, transplantation of HPCs is more complex due to their heterogeneity, the diversity of liver diseases and individual differences. It is necessary to further understand the regulation of HPC proliferation and differentiation before HPC transplantation can be applied as the gold standard treatment for liver diseases.
Long noncoding RNAs (lncRNAs) longer than 200 nucleotides (nt) are RNAs with noncoding functions. The lncRNAs are the most numerous, complex and the longest class of noncoding RNAs, and many of their functions remain to be revealed.10 Previous studies have reported that lncRNAs are not only involved in most cellular activities, including cell growth, apoptosis, differentiation, inflammatory response, and angiogenesis, but are also directly linked to the development of a diverse set of diseases.11, 12, 13 Recently, it has been pointed out that lncRNAs play an important role in the proliferation of hepatocytes and liver regeneration. Ruan et al. reported that lncRNA-Dreh could assist in the proliferation and migration of the HPC line WB-F344 during liver regeneration.14 Furthermore, Zhang et al. found that the downregulation of lncRNA metastasis-associated lung adenocarcinoma transcript 1 (lncRNA-MALAT1) enhanced the ligands of the transforming growth factor-β (TGF-β)/small mother against decapentaplegic signaling responses, and then induced the differentiation of pluripotent stem cells into hepatocytes.15 Deemed an oncogene, long noncoding RNAs/small nucleolar RNA host gene 12 (lncRNA SNHG12) was able to promote the development of a variety of tumors.16 Ruan et al. found that lncRNA SNHG12 accelerates the proliferation and migration of human osteosarcoma cells by boosting vascular actin gene expression.17 A study by Wang et al. revealed that the overexpression of lncRNA SNHG12 was capable of promoting cell proliferation and migration, as well as inhibiting apoptosis in triple-negative breast cancer.18 Lan et al. showed that lncRNA SNHG12 contributed to the development of hepatocellular carcinoma (HCC) by targeting microRNA-199a-5p.19 Additionally, that study reported that lncRNA SNHG12 was upregulated after 2/3 of hepatectomy in mice, and the overexpression of lncRNA SNHG12 activated the Wnt/β-catenin signaling pathway to promote liver regeneration.20 Those studies revealed the importance of lncRNA SNHG12 in the proliferation of hepatocytes and liver regeneration. However, it is still unclear whether lncRNA SNHG12 has the same function in HPCs. Therefore, this study aims to explore the effect of lncRNA SNHG12 on the proliferation of HPCs. When lncRNA SNHG12 is overexpressed or knocked down in the HPC cell line WB-F344 in vitro, the changes in cell function such as cell proliferation, migration and cell cycle progression can be observed.
Objectives
This study aims to explore the effect of lncRNA SNHG12 on the proliferation of HPCs, which may provide new ideas for improving HPC transplantation.
Materials and methods
Cell culture
The WB-F344 cells at the 3rd passage were purchased from the American Type Culture Collection (ATCC) cell bank (Manassas, USA). All cells used as test samples were within 15 passages. The WB-F344 cells were cultured in Dulbecco’s modified Eagle’s medium/Ham’s F-12 Nutrient Mixture (DMEM/F12; Gibco, Waltham, USA) containing 10% fetal bovine serum (FBS; Gibco) and 100 U/L penicillin/streptomycin (Solarbio, Beijing, China) at 37°C in an incubator with 5% CO2.
Cell transfection and grouping
When reaching the logarithmic growth phase, the cells were digested using 0.25% trypsin (Solarbio). Then, the cells were resuspended in culture medium and seeded into a 6-well plate at 4×104 cells/well. Empty vector pcDNA3.1 (NC vector), pcDNA3.1-SNHG12, negative short hairpin RNA (sh-NC), and SNHG12 shRNA (sh-SNHG12) were constructed and synthesized by RIBOBIO (Guangzhou, China). Next, the WB-F344 cells were transfected with the above fragments when they reached 70% confluence using the Lipofectamine™ 2000 kit (Thermo Fisher Scientific, Waltham, USA) according to the manufacturer’s instructions. The WB-F344 cells were grouped as follows: no treatment (sham group), NC vector (NC vector group), pcDNA3.1-SNHG12 (SNHG12 group), sh-NC (sh-NC group), sh-SNHG12 (sh-SNHG12 group), pcDNA3.1-SNHG12 and 10 μM salinomycin (MedChemExpress, Shanghai, China) (SNHG12+salinomycin group).
Real-time quantitative polymerase chain reaction
Total RNA was extracted from cells using the TRIzol reagent (Thermo Fisher Scientific). Then, RNA was reverse-transcribed into cDNA using a random primer reverse transcription kit (Thermo Fisher Scientific). The real-time quantitative polymerase chain reaction (qPCR) assays were performed using the SYBR GREEN qPCR kit (Takara, Shiga, Japan) according to the manufacturer’s instructions, with cDNA as the template and lncRNA SNHG12 primers (Table 1). The β-actin was adopted as an internal control, and 6 replicates were used in the experiment. The experimental data obtained through qPCR were analyzed using the 2−ΔΔCt method to calculate the relative expression of the target genes.
Cell Counting Kit-8 test
The WB-F344 cells were seeded into 96-well plates at 1×103 cells/well. After 24 h, the cells were transfected with lncRNA SNHG12 or treated with reagents. After serum starvation for 1 night, the medium was replaced and cells were cultured for 24 h, 48 h and 72 h. Cell Counting Kit-8 (CCK-8) cell dye solution (10 µL) (Beyotime Biotechnology Co., Ltd., Shanghai, China) was added to each well and plates were placed in an incubator for 1.5 h. The optical density (OD) at 450 nm was measured with a microplate reader (VL0000D2; Thermo Fisher Scientific). The proliferation rate of the cells was calculated from the absorbance values.
5-ethynyl-2’-deoxyuridine analysis
The treated WB-F344 cells were seeded in a 24-well plate. Then, the cells were stained using a 5-ethynyl-2’-deoxyuridine (EdU) staining kit (Thermo Fisher Scientific) and mounted with neutral resin. Fields (n = 6–10) were randomly selected for observation of cells under a fluorescence microscope (model FM-600; Shanghai Pudan Optical Instruments Co., Ltd., Shanghai, China). The number of positively stained cells (red) in each field was recorded. The EdU labeling rate (%) was calculated as the number of positive cells/total cell count × 100%.
Flow cytometry detection
The cells were cultured for 24 h and digested with trypsin after transfection. The cells were centrifuged at 800 rpm for 5 min at 4°C and washed twice with ice-cold sterile phosphate-buffered saline (PBS; Solarbio). Then, the cells were resuspended in 1 mL of ice-cold methanol with a volume fraction of 70% and fixed overnight at 4°C. After centrifugation (800 rpm, 4°C, 5 min), the cells were washed twice with ice-cold sterile PBS. Then, 500 μL PBS was adopted to resuspend cells, and propidium iodide (PI; 50 μg/mL) was added. Finally, the cells were incubated at room temperature for 15 min in the dark, and the cell cycle was analyzed with flow cytometry.
Transwell assay
After the transfection for 24 h, 2×104 cells were added in the upper chamber of a transwell, and 700 μL of medium containing 20% FBS was added to the lower chamber. The inserts were taken out after 12–24 h of culture at 37°C and 5% CO2. Next, the inserts were washed 3 times with PBS, fixed with 1% glutaraldehyde for 30 min, and then washed again with PBS. Subsequently, the inserts were dried, and the cells were incubated with 0.1% crystal violet for 12 h. Then, the cells were washed with PBS, dried and observed with optical microscopy to identify cells that penetrated to the reverse side of the transwell inserts. Then, 6–10 fields were randomly selected to observe and record the number of positive cells in each field. Eventually, 3 fields were chosen to be photographed and analyzed.
Albumin and alpha-fetoprotein level detection
After 48 h of transfection, the cells were digested with trypsin and seeded in culture plates. Subsequently, the cells were cultured with minimal medium in the absence of FBS. After 24 h, the supernatant was collected and centrifuged at 800 rpm for 5 min at 4°C. The levels of albumin (ALB) and alpha-fetoprotein (AFP) were detected using an ALB assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) and AFP assay kit (Nanjing Jiancheng Bioengineering Institute) in accordance with the manufacturer’s instructions.
Western blot assay
The total proteins were extracted from the treated cells using radioimmunoprecipitation assay (RIPA) buffer (Sigma-Aldrich, St. Louis, USA). The concentration of total proteins was determined using a bicinchoninic acid (BCA) kit (Thermo Fisher Scientific), and 25 μg of proteins were boiled and denatured, then combined with ×1 loading buffer. The proteins were separated using sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and transferred to polyvinylidene difluoride (PVDF) membranes (MilliporeSigma, Bedford, USA) using wettability conversion method. The membranes were blocked with 5% nonfat dry milk, and incubated with the primary antibody (Abcam, Waltham, USA) overnight at 4°C. Then, the membranes were washed 3 times, and the secondary antibody (Abcam) was added for 1 h. After washing 3 times, enhanced chemiluminescence (ECL) reagent (Sigma-Aldrich) was added to the membranes. A gel imaging system was utilized to collect images. Any visible protein bands were analyzed using ImageJ software (National Institutes of Health, Bethesda, USA) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as internal control to calculate the relative protein expression.
Statistical analyses
The statistical analysis was performed using IBM SPSS v. 22.0 software (IBM Corp., Armonk, USA) and GraphPad Prism 9 (GraphPad Software, San Diego, USA) for mapping. All data are presented as mean with 95% confidence interval (95% CI). All experiments were repeated independently 3 times. Data that passed normality (Shapiro–Wilk test) and homogeneity of variance tests (Brown–Forsythe test) (shown in Table 2, Table 3, Table 4, Table 5, Table 6, Table 7, Table 8) were compared using one-way analysis of variance (ANOVA) followed by the Tukey’s post hoc test in order to analyze the differences among multiple groups. A value of p < 0.05 indicated a significant difference.
Results
Overexpression of lncRNA SNHG12 promoted the proliferation of WB-F344 cells
To explore the effect of lncRNA SNHG12 on the proliferation of WB-F344 cells, we analyzed cell proliferation in each group using CCK-8 and EdU assays. Furthermore, qPCR showed that compared with the NC vector group, lncRNA SNHG12 expression was significantly increased in the SNHG12 group (p < 0.05; Table 2). Compared with the sh-NC group, the expression of lncRNA SNHG12 in the sh-SNHG12 group was significantly decreased (p < 0.05; Figure 1A, Table 2). These results revealed that lncRNA SNHG12 was successfully knocked down or overexpressed through transfection. Furthermore, the proliferation rate of WB-F344 cells in the SNHG12 group was higher than in the NC vector group (p < 0.05; Table 3), while the knockdown of lncRNA SNHG12 reduced the cell proliferation rate (p < 0.05; Figure 1B, Table 3). The EdU staining demonstrated that the overexpression of lncRNA SNHG12 increased, while the knockdown of lncRNA SNHG12 decreased the percentage of EdU-positive cells (p < 0.05; Figure 1C,D, Table 4). It suggested that the lncRNA SNHG12 expression level was positively correlated with the proliferation of WB-F334 cells.
Overexpression of lncRNA SNHG12 promoted the migration of WB-F344 cells
The results of the transwell assay showed that the migration ability of WB-F344 cells in the SNHG12 group was significantly increased compared with the NC vector group. When compared with the sh-NC group, the sh-SNHG12 group presented a notable decrease in the migratory ability of WB-F344 cells (Figure 2). This suggests that the overexpression of lncRNA SNHG12 promoted the migration of WB-F344 cells.
Overexpression of lncRNA SNHG12 expedited cell cycle progression of WB-F344 cells
Results of the cell cycle analysis revealed that when compared with the NC vector group, the number of cells in G0/G1 phase was significantly lower, while the number of cells in S phase was significantly higher in the SNHG12 group. In comparison with the sh-NC group, the sh-SNHG12 group showed an increase in the proportion of cells in G0/G1 phase and a significant decrease in the proportion of S phase cells (Figure 3), suggesting that lncRNA SNHG12 promoted S phase progression of WB-F334 cells.
Overexpression of lncRNA SNHG12 promoted the differentiation of WB-F344 cells into hepatocytes
The ALB and AFP kits were used to detect the effect of intervention with lncRNA SNHG12 expression on ALB and AFP synthesis in HPCs (Figure 4A,B). Compared with the NC vector group, ALB levels were increased and AFP levels were significantly decreased in the SNHG12 group. When compared with the sh-NC group, the sh-SNHG12 group displayed a lower ALB level and a higher AFP level (Figure 4A,B; Table 5, Table 6). These results revealed that WB-F344 cells showed an increase in ALB synthesis and a decrease in AFP synthesis after the overexpression of lncRNA SNHG12. The overexpression of lncRNA SNHG12 promoted the differentiation of WB-F344 cells into hepatocytes.
Overexpression of lncRNA SNHG12 activated the Wnt/β-catenin signaling pathway in WB-F344 cells
Studies have shown that the Wnt/β-catenin signaling pathway could mediate the regeneration of hepatocytes.21 We hypothesized that lncRNA SNHG12 stimulated the Wnt/β-catenin signaling pathway to participate in the proliferation, migration and differentiation of HPCs. To verify this, we used western blot to detect the expression of proteins related to the Wnt/β-catenin signaling pathway. The results showed that the expression of β-catenin, cyclin D1 and c-Myc protein significantly increased in the SNHG12 group compared with the NC vector group (p < 0.05; Table 7). When compared with the NC vector group, the expression of β-catenin, cyclin D1 and c-Myc protein was significantly decreased in the sh-SNHG12 group (p < 0.05; Figure 5A, Table 7).
Finally, WB-F344 cells were treated with salinomycin (a specific inhibitor of the Wnt/β-catenin pathway). The western blot results indicated that the expression of β-catenin, cyclin D1 and c-Myc protein were significantly increased in the SNHG12 group compared with the NC vector group. The expression level of β-catenin, cyclin D1 and c-Myc protein were further notably decreased in the SNHG12+salinomycin group compared with the SNHG12 group (p < 0.05; Figure 5B, Table 8). Hence, it was suggested that lncRNA SNHG12 was able to promote the proliferation of hepatocytes through activation of the Wnt/β-catenin pathway.
Discussion
Liver transplantation is the only effective therapy for critical liver diseases. However, because of the global shortage of donor organs, many patients do not have access to this therapy. Hepatic progenitor cell therapy is theorized to be a remarkable alternative to liver transplantation.22 However, HPC has presented unsatisfactory therapeutic effects when applied alone. Therefore, it is necessary to find some auxiliary factors to improve the efficacy of HPC transplantation. Studies have highlighted that the proliferative ability of HPC is positively correlated with the severity of liver injury. There are many factors that affect the activation, proliferation and differentiation of HPCs. Some cytokines or inflammatory factors that can regulate a variety of biological behaviors of HPCs have been found, including tumor necrosis factor alpha (TNF-α)-like weak inducer of apoptosis, hepatocyte growth factor, epidermal growth factor, and interleukin 6 (IL-6).23, 24
In addition to their profound effect on liver transplantation, lncRNAs have been found to play an important role in a variety of biological processes. Xie et al. discovered that long noncoding small nucleolar RNA host gene 7 could bind and inhibit miR-29b expression in hepatic stellate cells and affect DNA-methyltransferase 3A expression. In that case, the activation, autophagy and proliferation of hepatic stellate cells could be impacted.25 Liao et al. highlighted that the lncRNA G protein-coupled receptor 137b-pseudogene (lncRNA Gpr137b-ps) could affect the activation of C-X-C motif chemokine ligand 14 (CXCL14) by targeting and regulating the expression of miR-200a-3p, and thereby activating hepatic stellate cells.26 Zhang et al. reported that the upregulation of liver fibrosis-associated lncRNA1 expression in the liver promoted the transcription of TGF-β, some mothers against decapentaplegic homolog 2 which transmits the activation signal from TGF-β receptors at the plasma membrane to transcriptional regulators in the nucleus, mothers against decapentaplegic homolog 3, and neurogenic locus notch homolog protein 2 and 3-induced TGFβ and notch pathway activation, as well as inhibited hepatic stellate cell activation.27 However, there are few relevant studies on lncRNAs in HPCs. In our study, we found that lncRNA SNHG12 possessed several functions, including facilitating the proliferation, migration and cell cycle progression of WB-F344. Moreover, lncRNA SNHG12 was effective in upregulating ALB and downregulating AFP expression in cells. Alpha-fetoprotein is a glycoprotein derived from embryonic endodermal cells. Despite the high level in fetal serum, AFP content will gradually decrease to adult levels after birth. Therefore, AFP is often applied as a marker of HPC.28 Albumin, on the other hand, is often considered a marker of mature hepatocytes.29 It can thus be seen that lncRNA SNHG12 is conducive to the proliferation, migration, cell cycle progression, and differentiation of HPCs.
The Wnt/β-catenin not only is a signaling pathway for stem cell differentiation, but also plays a role in cell survival, proliferation, embryonic patterning, organogenesis, differentiation, cell migration, and polarity.21 Moreover, Wnt/β-catenin signaling regulates many biological processes in the development, homeostasis, metabolism, regeneration, and carcinogenesis of the liver.30 It has been shown that the downregulation of Wnt/β-catenin signaling activity can assist the differentiation of umbilical cord stem cells to hepatocytes.31 In a previous study, lncRNAs have been found to regulate the activation of the Wnt/β-catenin signaling pathway. Song et al. claimed that lncRNA AWPPH, promoted the proliferation and inhibited the apoptosis of non-small cell lung cancer cells by activating the Wnt/β-catenin signaling pathway.32 Liu et al. concluded that the upregulation of lncRNA nuclear-enriched abundant transcript 1 could promote colon cancer progression by sponging miR-486-5p and activating nuclear receptor subfamily 4 group A member 1/Wnt/β-catenin pathway.33 While Zhao et al. summarized that lncRNA SMAD5-AS1, as a competitive endogenous RNA, could increase adenomatous polyposis coli (APC) expression by stimulating miR-135b-5p and inhibit the proliferation of diffuse large B-cell lymphoma by the upregulation of the Wnt/β-catenin pathway.34 The above studies indicate that lncRNA could regulate Wnt/β-catenin signaling though a variety of methods, and produce various results. In our study, lncRNA SNHG12 was shown to be able to activate Wnt/β-catenin signaling. Several lncRNAs have been determined to be involved in Wnt/β-catenin signaling in order to elevate the proliferation of hepatocytes, including lncRNA liver regeneration 1, lncRNA transcription factor 7 and lncRNA lnc-β-Catm, which is highly expressed in human HCC tumors and liver cancer stem cells.35, 36 Therefore, it can be suggested that lncRNA SNHG12 exerts its biological function by activating the Wnt/β-catenin signaling pathway.
Limitations
There are limitations to this study. The lncRNAs have multiple targets and the way lncRNA SNHG12 regulates Wnt/β-catenin signaling needs further exploration. In addition, the involvement of lncRNA SNHG12 in the regulation of hepatocyte proliferation was only confirmed through in vitro cellular experiments; therefore, in vivo animal experiments need to be performed. Finally, because of the very small sample sizes used in the ANOVA, it is more difficult to reliably verify the test assumptions, and our results need to be considered with caution.
Conclusions
In summary, the upregulation of lncRNA SNHG12 expression in WB-F344 cells activated the Wnt/β-catenin signaling pathway, and promoted cell proliferation, migration and cell cycle progression. However, lncRNAs have been shown to have multiple targets. Further studies are required to thoroughly elucidate the mechanism of action of lncRNA SNHG12 in order to provide a solid basis for clinical application.
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)